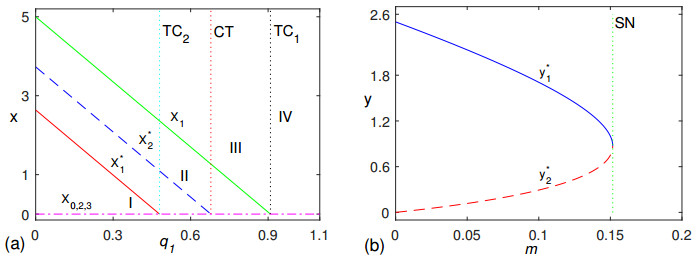
In this paper, in order to explore the inhibition mechanism of algicidal bacteria on algae, we constructed an aquatic amensalism model with non-selective harvesting and Allee effect. Mathematical works mainly gave some critical conditions to guarantee the existence and stability of equilibrium points, and derived some threshold conditions for saddle-node bifurcation and transcritical bifurcation. Numerical simulation works mainly revealed that non-selective harvesting played an important role in amensalism dynamic relationship. Meanwhile, we proposed some biological explanations for transcritical bifurcation and saddle-node bifurcation from the aspect of algicidal bacteria controlling algae. Finally, all these results were expected to be useful in studying dynamical behaviors of aquatic amensalism ecosystems and biological algae controlling technology.
Citation: Huanyi Liu, Hengguo Yu, Chuanjun Dai, Zengling Ma, Qi Wang, Min Zhao. Dynamical analysis of an aquatic amensalism model with non-selective harvesting and Allee effect[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8857-8882. doi: 10.3934/mbe.2021437
In this paper, in order to explore the inhibition mechanism of algicidal bacteria on algae, we constructed an aquatic amensalism model with non-selective harvesting and Allee effect. Mathematical works mainly gave some critical conditions to guarantee the existence and stability of equilibrium points, and derived some threshold conditions for saddle-node bifurcation and transcritical bifurcation. Numerical simulation works mainly revealed that non-selective harvesting played an important role in amensalism dynamic relationship. Meanwhile, we proposed some biological explanations for transcritical bifurcation and saddle-node bifurcation from the aspect of algicidal bacteria controlling algae. Finally, all these results were expected to be useful in studying dynamical behaviors of aquatic amensalism ecosystems and biological algae controlling technology.

| [1] |
T. F. Steppe, J. B. Olson, H. W. Paerl, Consortial N2 fixation: A strategy for meeting nitrogen requirements of marine and terrestrial cyanobacterial mats, FEMS. Microbiol. Ecol., 21 (1996), 149–156. doi: 10.1111/j.1574-6941.1996.tb00342.x

|
| [2] |
A. Dakhama, J. Noue, M. C. Lavoie, Isolation and identification of antialgal substances produced by Pseudomonas aeruginosa, J. Appl. Phycol., 5 (1993), 297–306. doi: 10.1007/BF02186232
|
| [3] | Y. Kawano, Y. Nagawa, H. Nakanishi, H. Nakajima, M. Matsuo, T. Higashihara, Production of thiotropocin by a marine bacterium Caulobacter sp and its antimicroalgal activities, J. Mar. Biotechnol., 5 (1997), 225–229. |
| [4] | C. Q. Xiao, H. Jiang, K. Cheng, Y. J. Zhao, Selection of algae lysing actinomycetes AN02 and optimization of its cultural conditions, J. Microbiol., 27 (2007), 11–14. |
| [5] |
H. J. Choi, B. H. Baik, J. D. Kim, M. S. Han, Streptomyces neyagawaensis as a control for the hazardous biomass of Microcystis aeruginosa (Cyanobacteria) in eutrophic freshwaters, Biol. Control., 33 (2005), 335–343. doi: 10.1016/j.biocontrol.2005.03.007
|
| [6] |
Y. F. Yang, X. J. Hu, J. Zhang, Y. X. Gong, Community level physiological study of algicidal bacteria in the phycospheres of Skeletonema costatum and Scrippsiella trochoidea, Harmful. Algae., 28 (2013), 88–96. doi: 10.1016/j.hal.2013.05.015
|
| [7] | E. Odum, Fundamentals of ecology, Saunders, Philadelphia, 1953. |
| [8] | J. Li, Ecology, Science Press, 2014. |
| [9] | G. C. Sun, Qualitative analysis on two populations amensalism model, J. Jiamusi Univ., 21 (2003), 283–286. |
| [10] | Z. F. Zhu, Q. L. Chen, Mathematic analysis on commensalism Lotka-Volterra model of populations, J. Jixi. Univ., 8 (2008), 100–101. |
| [11] | F. D. Chen, W. X. He, R. Y. Han, On discrete amensalism model of Lotka-Volterra, J. Beihua. Univ., 16 (2015), 141–144. |
| [12] |
H. H. Xiong, B. B. Wang, H. L. Zhang, Stability analysis on the dynamic model of fish swarm amensalism, Adv. Appl. Math., 5 (2016), 255–261. doi: 10.12677/AAM.2016.52032
|
| [13] | R. X. Wu, A two species amensalism model with non-monotonic functional response, Commun. Math. Biol. Neurosci., 2016 (2016), 19. |
| [14] |
X. Y. Guan, F. D. Chen, Dynamical analysis of a two species amensalism model with Beddington-DeAngelis functional response and Allee effect on the second species, Nonlinear. Anal. RWA., 48 (2019), 71–93. doi: 10.1016/j.nonrwa.2019.01.002
|
| [15] | L. C. Zhao, Q. L. Zhang, Q. C. Yang, The induction control of a three-species model with partial harm relations, J. Biomath., 20 (2005), 37–42. |
| [16] | R. X.Wu, L. Li, Q. F. Lin, A Holling type commensal symbiosis model involving Allee effect, Commun. Math. Biol. Neurosci., 2018 (2018), 6. |
| [17] |
Q. Q. Su, F. D. Chen, The influence of partial closure for the populations to a non-selective harvesting Lotka-Volterra discrete amensalism model, Adv. Differ. Equ-Ny., 2019 (2019), 281. doi: 10.1186/s13662-019-2209-y
|
| [18] | R. Y. Han, Y. L. Xue, L. Y. Yang, F. D. Chen, On the existence of positive periodic solution of a Lotka-Volterra amensalism model, J. Longyan. Univ., 33 (2015), 22–26. |
| [19] |
Z. Wei, Y. H. Xia, T. H. Zhang, Stability and bifurcation analysis of an amensalism model with weak Allee effect, Qual. Theor. Dyn. Syst., 19 (2020), 23. doi: 10.1007/s12346-020-00341-0
|
| [20] | F. Courchamp, L. Berec, J. Gascoigne, Allee effects in ecology and conservation, Oxford University Press, New York, 2008. |
| [21] | W. C. Allee, Animal aggragations, a study in general sociology, University of Chicago Press, Chicago, 1931. |
| [22] |
P. A. Stephens, W. J. Sutherland, Consequences of the Allee effect for behavior, ecology and conservation, Trends. Ecol. Evol., 14 (1999), 401–405. doi: 10.1016/S0169-5347(99)01684-5
|
| [23] |
F. Courchamp, T. Clutton-Brock, B. Grenfell, Inverse density dependence and the Allee effect, Trends. Ecol. Evol., 14 (1999), 405–410. doi: 10.1016/S0169-5347(99)01683-3
|
| [24] |
P. A. Stephens, W. J. Sutherland, R. P. Freckleton, What is the Allee effect?, Oikos, 87 (1999), 185–190. doi: 10.2307/3547011
|
| [25] |
M. Liermann, R. Hilborn, Depensation: Evidence, models and implication, Fish Fish., 2 (2001), 33–58. doi: 10.1046/j.1467-2979.2001.00029.x
|
| [26] | G. P. Stamou, M. D. Asikidis, The effect of density on the demographic parameters of two oribatid mites, Rev. Ecol. Biol. Sology., 26 (1989), 321–330. |
| [27] | F. Courchamp, L. Berec, J. Gascoigne, Allee effects in ecology and conservation, Oxford University Press, New York, 2008. |
| [28] |
C. Rebelo, C. Soresina, Coexistence in seasonally varying predator-prey systems with Allee effect, Nonlinear Anal. RWA, 55 (2020), 103140. doi: 10.1016/j.nonrwa.2020.103140
|
| [29] |
U. Kumar, P. S. Mandal, E. Venturino, Impact of Allee effect on an eco-epidemiological system, Ecol. Complex., 42 (2020), 100828. doi: 10.1016/j.ecocom.2020.100828
|
| [30] |
D. Sen, S. Ghorai, S. Sharma, M. Banerjee, Allee effect in prey growth reduces the dynamical complexity in prey-predator model with generalist predator, Appl. Math. Model., 91 (2021), 768–790. doi: 10.1016/j.apm.2020.09.046
|
| [31] |
D. Y. Bai, Y. Kang, S. G. Ruan, L. S. Wang, Dynamics of an intraguild predator food web model with strong Allee effect in the basal prey, Nonlinear Anal. RWA, 58 (2021), 103206. doi: 10.1016/j.nonrwa.2020.103206
|
| [32] |
J. P. Tripathi, P. S. Mandal, A. Poonia, V. P. Bajiya, A widespread interaction between generalist and specialist enemies: The role of intraguild predation and Allee effect, Appl. Math. Model., 89 (2021), 105–135. doi: 10.1016/j.apm.2020.06.074
|
| [33] | P. G. Brown, P. Timmerman, Ecological Economics for the Anthropocene: An Emerging Paradigm, Columbia University Press, United States, 2015. |
| [34] |
T. K. Ang, H. M. Safuan, Dynamical behaviors and optimal harvesting of an intraguild prey-predator fishery model with Michaelis-Menten type predator harvesting, BioSystems, 202 (2021), 104357. doi: 10.1016/j.biosystems.2021.104357
|
| [35] |
C. W. Clark, Mathematical models in the economics of renewable resources, SIAM Rev., 21 (1979), 81–99. doi: 10.1137/1021006
|
| [36] | C. W. Clark, M. Mangel, Aggregation and fishery dynamics: A theoretical study of schooling and the purse seine tuna fisheries, Fish. B-Noaa., 77 (1979), 317–337. |
| [37] |
T. Das, R. N. Mukherjee, K. S. Chaudhuri, Harvesting of a prey-predator fishery in the presence of toxicity, Appl. Math. Model., 33 (2009), 2282–2292. doi: 10.1016/j.apm.2008.06.008
|
| [38] |
T. Das, R. N. Mukherjee, K. S. Chaudhuri, Bio-economic harvesting of a prey-predator fishery, J. Biol. Dynam., 3 (2009), 447–462. doi: 10.1080/17513750802560346
|
| [39] |
X. X. Liu, Q. D. Huang, Analysis of optimal harvesting of a predator-prey model with Holling type IV functional response, Ecol. Complex., 42 (2020), 100816. doi: 10.1016/j.ecocom.2020.100816
|
| [40] |
E. Bellier, B. E. Sether, S. Engen, Sustainable strategies for harvesting predators and prey in a fluctuating environment, Ecol. Model., 440 (2021), 109350. doi: 10.1016/j.ecolmodel.2020.109350
|
| [41] |
J. Datta, D. Jana, R. K. Upadhyay, Bifurcation and bio-economic analysis of a prey-generalist predator model with Holling type IV functional response and nonlinear age-selective prey harvesting, Chaos Soliton. Fract., 122 (2019), 229–235. doi: 10.1016/j.chaos.2019.02.010
|
| [42] |
M. Li, B. S. Chen, H. W. Ye, A bioeconomic differential algebraic predator-prey model with nonlinear prey harvesting, Appl. Math. Model., 42 (2017), 17–28. doi: 10.1016/j.apm.2016.09.029
|
| [43] | R. P. Gupta, P. Chandra, M. Banerjee, Dynamical complexity of a prey-predator model with nonlinear predator harvesting, Discrete. Cont. Dyn-B., 20 (2015), 423–443. |
| [44] | L. N. Guin, S. Pal, S. Chakravarty, S. Djilali, Pattern dynamics of a reaction-diffusion predator-prey system with both refuge and harvesting, Int. J. Biomath., 14 (2020), 2050084. |
| [45] | L. N. Guin, R. Murmu, H. Baek, K. H. Kim, Dynamical analysis of a Beddington-DeAngelis interacting species system with prey harvesting, Math. Probl. Eng., 2020 (2020), 22. |
| [46] |
L. N. Guin, S. Acharya, Dynamic behaviour of a reaction-diffusion predator-prey model with both refuge and harvesting, Nonlinear Dynam., 88 (2017), 1–33. doi: 10.1007/s11071-016-3227-x
|
| [47] |
H. G. Yu, M. Zhao, R. P. Agarwal, Stability and dynamics analysis of time delayed eutrophication ecological model based upon the Zeya reservoir, Math. Comput. Simulat., 97 (2014), 53–67. doi: 10.1016/j.matcom.2013.06.008
|
| [48] | L. V. Bertalanffy, Theoretische biologise, Berlin, 1932. |
| [49] |
H. G. Yu, M. Zhao, Q, Wang, R. P. Agarwal, A focus on long-run sustainability of an impulsive switched eutrophication controlling system based upon the Zeya reservoir, J. Franklin Inst., 351 (2014), 487–499. doi: 10.1016/j.jfranklin.2013.08.025
|
| [50] | H. Malchow, S. Petrovskii, A. Medvinsky, Pattern formation in models of plankton dynamics: A synthesis, Oceanol. Acta., 24 (2011), 479–487. |
| [51] |
D. Balram, K. Ankit, P. M. Atasi, Global stability and Hopf-bifurcation of prey-predator system with two discrete delays including habitat complexity and prey refuge, Commun. Nonlinear Sci. Numer. Simulat., 67 (2019), 528–554. doi: 10.1016/j.cnsns.2018.07.019
|
| [52] |
L. N. Guin, E. Das, M. Sambath, Pattern formation scenario via Turing instability in interacting reaction-diffusion systems with both refuge and nonlinear harvesting, J. Appl. Nonl. Dyn., 9 (2020), 1–21. doi: 10.5890/JAND.2020.03.001
|
| [53] |
L. N. Guin, H. Baek, Comparative study between prey-dependent and ratio-dependent predator-prey models relating to patterning phenomenon, Math. Comput. Simulat., 146 (2018), 100–117. doi: 10.1016/j.matcom.2017.10.015
|
| [54] |
L. N. Guin, B. Mondal, S. Chakravarty, Spatiotemporal patterns of a pursuit-evasion generalist predator-prey model with prey harvesting, J. Appl. Nonl. Dyn., 7 (2018), 165–177. doi: 10.5890/JAND.2018.06.005
|
| [55] | S. T. Wang, H. G. Yu, Complexity anaylsis of a modified predator-prey system with Beddington-DeAngelis functional response and Allee-like effect on predator, Discrete Dyn. Na. Soc., 2021 (2021), 1–18. |
| [56] |
X. X. Li, H. G. Yu, C. J. Dai, Z. L. Ma, Q. Wang, M. Zhao, Bifurcation analysis of a new aquatic ecological model with aggregation effect, Math. Comput. Simulat., 190 (2021), 75–96. doi: 10.1016/j.matcom.2021.05.015
|
| [57] | R. J. Han, L. N. Guin, B. X. Dai, Cross-diffusion-driven pattern formation and selection in a modified Leslie-Gower predator-prey model with fear effect, J. Biol. Syst., 28 (2020), 1–38. |
| [58] |
R. J. Han, L. N. Guin, B. X. Dai, Consequences of refuge and diffusion in a spatiotemporal predator-prey model, Nonlinear Anal. RWA., 60 (2021), 103311. doi: 10.1016/j.nonrwa.2021.103311
|
| [59] | H. J. Guo, W. Zhang, X. M. Zhang, L. Q. Mao, W. Y. Zang, Process of alge-lysis and chlorophyll degradation kinetics of Microbacterium oleivoran bacteria, Environ. Chem., 190 (2021), 75-96. |
| [60] | S. E. Jorgensen, G. Bendoricchio, Fundamentals of Ecological Modelling, Elsevier BV, The Netherlands, 2001. |
| [61] |
D. P. Hu, H. J. Cao, Stability and bifurcation analysis in a predator-prey system with Michaelis-Menten type predator harvesting, Nonlinear Anal. RWA., 33 (2017), 58–82. doi: 10.1016/j.nonrwa.2016.05.010
|
| [62] | Z. F. Zhang, W. Z. Huang, Z. X. Dong, Qualitative Theory of Differential Equation, Science Press, Beijing, 1992. |
| [63] | L. Perko, Differential Equations and Dynamical Systems, Springer, New York, 2001. |




Figures(5) / Tables(1)

Huanyi Liu, Hengguo Yu, Chuanjun Dai, Zengling Ma, Qi Wang, Min Zhao. Dynamical analysis of an aquatic amensalism model with non-selective harvesting and Allee effect[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8857-8882. doi: 10.3934/mbe.2021437







 DownLoad:
DownLoad: