In the paper, a Leslie-Gower predator-prey system with harvesting and fear effect is considered. The existence and stability of all possible equilibrium points are analyzed. The bifurcation dynamic behavior at key equilibrium points is investigated to explore the intrinsic driving mechanisms of population interaction modes. It is shown that the system undergoes various bifurcations, including transcritical, saddle-node, Hopf and Bogdanov-Takens bifurcations. The numerical simulation results show that harvesting and fear effect can seriously affect the dynamic evolution trend and coexistence mode. Furthermore, it is particularly worth pointing out that harvesting not only drives changes in population coexistence mode, but also has a certain degree delay. Finally, it is anticipated that these research results will be beneficial for the vigorous development of predator-prey system.
Citation: Rongjie Yu, Hengguo Yu, Chuanjun Dai, Zengling Ma, Qi Wang, Min Zhao. Bifurcation analysis of Leslie-Gower predator-prey system with harvesting and fear effect[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18267-18300. doi: 10.3934/mbe.2023812
In the paper, a Leslie-Gower predator-prey system with harvesting and fear effect is considered. The existence and stability of all possible equilibrium points are analyzed. The bifurcation dynamic behavior at key equilibrium points is investigated to explore the intrinsic driving mechanisms of population interaction modes. It is shown that the system undergoes various bifurcations, including transcritical, saddle-node, Hopf and Bogdanov-Takens bifurcations. The numerical simulation results show that harvesting and fear effect can seriously affect the dynamic evolution trend and coexistence mode. Furthermore, it is particularly worth pointing out that harvesting not only drives changes in population coexistence mode, but also has a certain degree delay. Finally, it is anticipated that these research results will be beneficial for the vigorous development of predator-prey system.

| [1] |
A. J. Lotka, Elements of physical biology, Nature, 461 (1925). https://doi.org/10.1038/116461b0 doi: 10.1038/116461b0

|
| [2] |
V. Volterra, Fluctuations in the abundance of a species considered mathematically, Nature, 1926 (1926), 558–560. https://doi.org/10.1038/118558a0 doi: 10.1038/118558a0
|
| [3] |
S. B. Hsu, T. W. Huang, Global stability for a class of predator-prey systems, SIAM J. Appl. Math, 55 (1995), 763–783. https://doi.org/10.1137/S0036139993253201 doi: 10.1137/S0036139993253201
|
| [4] |
D. M. Xiao, H. P. Zhu, Multiple focus and Hopf bifurcation in a predator-prey system with nonmonotonic functional response, SIAM J. Appl. Math, 66 (2006), 802–819. https://doi.org/10.1137/050623449 doi: 10.1137/050623449
|
| [5] |
Y. Lamontagne, C. Coutu, C. Rousseau, Bifurcation analysis of a predator-prey system with generalised Holling type Ⅲ functional response, J. Dynam. Differ. Equations, 20 (2008), 535–571. https://doi.org/10.1007/s10884-008-9102-9 doi: 10.1007/s10884-008-9102-9
|
| [6] |
D. M. Xiao, K. F. Zhang, Multiple bifurcations of a predator-prey system, Discrete Contin. Dynam. Syst. Ser. Ser. B, 8 (2007), 417–433. https://doi.org/10.3934/dcdsb.2007.8.417 doi: 10.3934/dcdsb.2007.8.417
|
| [7] |
R. M. Etoua, C. Rousseau, Bifurcation analysis of a generalized Gause model with prey harvesting and a generalized Holling response function of type Ⅲ, J. Differ. Equations, 249 (2010), 2316–2356. https://doi.org/10.1016/j.jde.2010.06.021 doi: 10.1016/j.jde.2010.06.021
|
| [8] |
L. L. Cai, G. T. Chen, D. M. Xiao, Multiparametric bifurcations of an epidemiological model with strong Allee effect, J. Math. Biol., 67 (2013), 185–215. https://doi.org/10.1007/s00285-012-0546-5 doi: 10.1007/s00285-012-0546-5
|
| [9] |
P. H. Leslie, Some further notes on the use of matrices in population mathematics, Biometrika, 35 (1948), 213–245. https://doi.org/10.2307/2332342 doi: 10.2307/2332342
|
| [10] |
P. H. Leslie, A stochastic model for studying the properties of certain biological systems by numerical methods, Biometrika, 45 (1958), 16–31. https://doi.org/10.1093/biomet/45.1-2.16 doi: 10.1093/biomet/45.1-2.16
|
| [11] |
M. A. Aziz-Alaoui, M. D. Okiye, Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type Ⅱ schemes, Appl. Math. Lett., 16 (2003), 1069–1075. https://doi.org/10.1016/S0893-9659(03)90096-6 doi: 10.1016/S0893-9659(03)90096-6
|
| [12] |
D. P. Hu, H. J. Cao, Stability and bifurcation analysis in a predator-prey system with Michaelis-Menten type predator harvesting, Nonlinear Anal. RWA, 33 (2017), 58–82. https://doi.org/10.1016/j.nonrwa.2016.05.010 doi: 10.1016/j.nonrwa.2016.05.010
|
| [13] |
M. Liu, Dynamics of a stochastic regime-switching predator-prey model with modified Leslie-Gower Holling-type Ⅱ schemes and prey harvesting, Nonlinear Dyn., 96 (2019), 417–442. https://doi.org/10.1007/s11071-019-04797-x doi: 10.1007/s11071-019-04797-x
|
| [14] |
Y. L. Li, D. M. Xiao, Bifurcations of a predator-prey system of Holling and Leslie types, Chaos Solitons Fractals, 34 (2007), 606–620. https://doi.org/10.1016/j.chaos.2006.03.068 doi: 10.1016/j.chaos.2006.03.068
|
| [15] |
Z. C. Shang, Y. H. Qiao, Bifurcation analysis of a Leslie-type predator-prey system with simplified Holling type Ⅳ functional response and strong Allee effect on prey, Nonlinear Anal. RWA, 64 (2022), 103453. https://doi.org/10.1016/j.nonrwa.2021.103453 doi: 10.1016/j.nonrwa.2021.103453
|
| [16] |
A. Arsie, C. Kottegoda, C. H. Shan, A predator-prey system with generalized Holling type Ⅳ functional response and Allee effects in prey, J. Differ. Equations, 309 (2022), 704–740. https://doi.org/10.1016/j.jde.2021.11.041 doi: 10.1016/j.jde.2021.11.041
|
| [17] |
Y. J. Li, M. X. He, Z. Li, Dynamics of a ratio-dependent Leslie-Gower predator-prey model with Allee effect and fear effect, Math. Comput. Simul., 201 (2022), 417–439. https://doi.org/10.1016/j.matcom.2022.05.017 doi: 10.1016/j.matcom.2022.05.017
|
| [18] |
J. C. Huang, Y. J. Gong, S. G. Ruan, Bifurcation analysis in a predator-prey model with constant-yield predator harvesting, Discrete Contin. Dyn. Syst. Ser. B, 18 (2013), 2101–2121. https://doi.org/10.3934/dcdsb.2013.18.2101 doi: 10.3934/dcdsb.2013.18.2101
|
| [19] |
J. Wang, Y. L. Cai, S. M. Fu, W. M. Wang, The effect of the fear factor on the dynamics of a predator-prey model incorporating the prey refuge, Chaos, 29 (2019), 083109. https://doi.org/10.1063/1.5111121 doi: 10.1063/1.5111121
|
| [20] | Z. F. Zhang, T. R. Ding, W. Z. Huang, Z. X. Dong, Qualitative Theory of Differential Equation, Science Press, 1992. |
| [21] |
J. Chen, J. C. Huang, S. G. Ruan, J. H. Wang, Bifurcations of invariant tori in predator-prey models with seasonal prey harvesting, SIAM J. Appl. Math., 73 (2013), 1876–1905. https://doi.org/10.1137/120895858 doi: 10.1137/120895858
|
| [22] |
J. C. Huang, Y. J. Gong, J. Chen, Multiple bifurcation in a predator-prey system of Holling and Leslie type with constant-yield prey harvesting, Int. J. Bifur. Chaos, 23 (2013), 1350164. https://doi.org/10.1142/S0218127413501642 doi: 10.1142/S0218127413501642
|
| [23] | L. Perko, Differential Equations and Dynamical Systems, Springer, 2001. |
| [24] |
L. Y. Zanette, A. F. White, M. C. Allen, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. https://doi.org/10.1126/science.1210908 doi: 10.1126/science.1210908
|
| [25] |
K. H. Elliott, G. S. Betini, D. R. Norris, Fear creates an Allee effect: experimental evidence from seasonal populations, Proc. R. Soc. B: Biol Sci, 284 (2017), 1950195. https://doi.org/10.1098/rspb.2017.0878 doi: 10.1098/rspb.2017.0878
|
| [26] |
X. Y. Wang, L. Zanette, X. F. Zou, Modelling the fear effect in predator-prey interactions, J. Math. Biol., 73 (2016), 1179–1204. https://doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [27] |
S. K. Sasmal, Population dynamics with multiple Allee effects induced by fear factors-A mathematical study on prey-predator interactions, Appl. Math. Model., 64 (2018), 1–14. https://doi.org/10.1016/j.apm.2018.07.021 doi: 10.1016/j.apm.2018.07.021
|
| [28] |
S. Pal, N. Pal, S. Samanta, J. Chattopadhyay, Effect of hunting cooperation and fear in a predator-prey model, Ecol. Complex, 39 (2019), 100770. https://doi.org/10.1016/j.ecocom.2019.100770 doi: 10.1016/j.ecocom.2019.100770
|
| [29] |
S. Pal, N. Pal, S. Samanta, J. Chattopadhyay, Fear effect in prey and hunting cooperation among predators in a Leslie-Gower model, Math. Biosci. Eng., 16 (2019), 5146–5179. https://doi.org/10.3934/mbe.2019258 doi: 10.3934/mbe.2019258
|
| [30] |
P. Panday, N. Pal, S. Samanta, J. Chattopadhyay, Stability and bifurcation analysis of a three-species food chain model with fear, Int. J. Bifurc. Chaos, 28 (2018), 1850009. https://doi.org/10.1142/S0218127418500098 doi: 10.1142/S0218127418500098
|
| [31] |
T. Qiao, Y. L. Cai, S. M. Fu, W. M. Wang, Stability and Hopf bifurcation in a predator-prey model with the cost of anti-predator behaviors, Int. J. Bifurc. Chaos, 29 (2019), 1950185. https://doi.org/10.1142/S0218127419501852 doi: 10.1142/S0218127419501852
|
| [32] |
K. Sarkar, S. Khajanchi, Impact of fear effect on the growth of prey in a predator-prey interaction model, Ecol. Complex, 42 (2020), 100826. https://doi.org/10.1016/j.ecocom.2020.100826 doi: 10.1016/j.ecocom.2020.100826
|
| [33] |
V. Tiwari, J. P. Tripathi, S. Mishra, R. K. Upadhyay, Modeling the fear effect and stability of non-equilibrium patterns in mutually interfering predator-prey systems, Appl. Math. Comput., 371 (2020), 124948. https://doi.org/10.1016/j.amc.2019.124948 doi: 10.1016/j.amc.2019.124948
|
| [34] |
X. Y. Wang, X. F. Zou, Modeling the fear effect in predator-prey interactions with adaptive avoidance of predators, Bull. Math. Biol., 79 (2017), 1325–1359. https://doi.org/10.1007/s11538-017-0287-0 doi: 10.1007/s11538-017-0287-0
|
| [35] |
H. S. Zhang, Y. L. Cai, S. M. Fu, W. M. Wang, Impact of the fear effect in a prey-predator model incorporating a prey refuge, Appl. Math. Comput., 356 (2019), 328–337. https://doi.org/10.1016/j.amc.2019.03.034 doi: 10.1016/j.amc.2019.03.034
|
| [36] |
X. B. Zhang, Q. An, L. Wang, Spatiotemporal dynamics of a delayed diffusive ratio-dependent predator-prey model with fear effect, Nonlinear Dyn., 105 (2021), 3775–3790. https://doi.org/10.1007/s11071-021-06780-x doi: 10.1007/s11071-021-06780-x
|
| [37] |
P. P. Cong, M. Fan, X. F. Zou, Dynamics of a three-species food chain model with fear effect, Commun. Nonlinear Sci. Numer. Simul., 99 (2021), 105809. https://doi.org/10.1016/j.cnsns.2021.105809 doi: 10.1016/j.cnsns.2021.105809
|
| [38] |
X. Q. Wang, Y. P. Tan, Y. L. Cai, W. M. Wang, Impact of the fear effect on the stability and bifurcation of a Leslie-Gower predator-prey model, Int. J. Bifurc. Chaos, 30 (2020), 2050210. https://doi.org/10.1142/S0218127420502107 doi: 10.1142/S0218127420502107
|
| [39] |
X. Y. Wang, X. F. Zou, Pattern formation of a predator-prey model with the cost of anti-predator behaviors, Math. Biosci. Eng., 15 (2018), 775–805. https://doi.org/10.3934/mbe.2018035 doi: 10.3934/mbe.2018035
|
| [40] |
R. J. Han, L. N. Guin, B. X. Dai, Cross-diffusion-driven pattern formation and selection in a modified leslie-gower predator-prey model with fear effect, J. Biol. Syst., 28 (2020), 27–64. https://doi.org/10.1142/S0218339020500023 doi: 10.1142/S0218339020500023
|
| [41] |
S. Li, S. L. Yuan, Z. Jin, H. Wang, Bifurcation analysis in a diffusive predator-prey model with spatial memory of prey, Allee effect and maturation delay of predator, J. Differ. Equations, 357 (2023), 32–63. https://doi.org/10.1016/j.jde.2023.02.009 doi: 10.1016/j.jde.2023.02.009
|
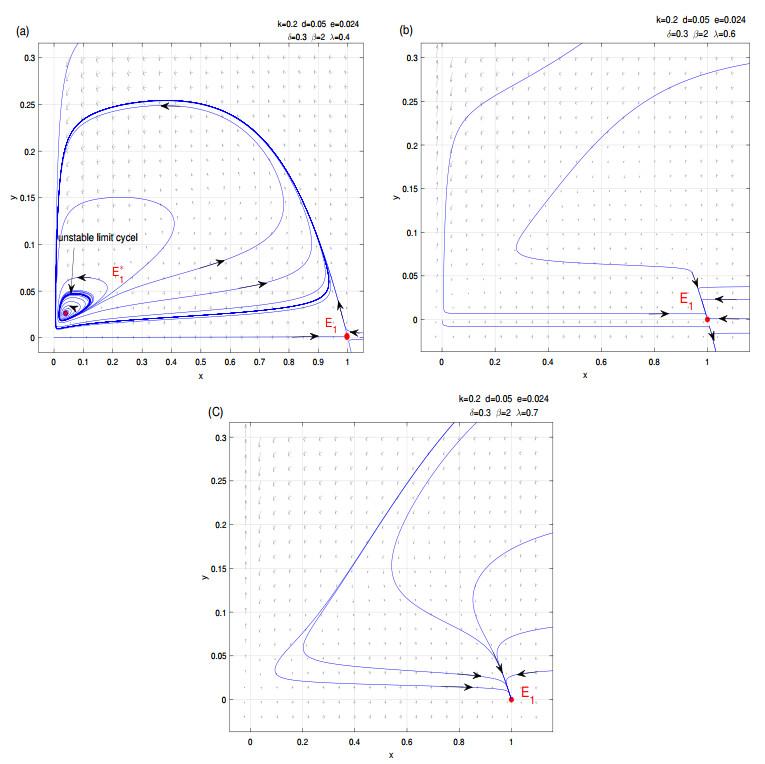







Figures(8)

Rongjie Yu, Hengguo Yu, Chuanjun Dai, Zengling Ma, Qi Wang, Min Zhao. Bifurcation analysis of Leslie-Gower predator-prey system with harvesting and fear effect[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18267-18300. doi: 10.3934/mbe.2023812







 DownLoad:
DownLoad: