Citation: Feng Chen, Xuqing Zhu, Jing Zheng, Tingting Xu, Kuan Wu, Chuhui Ru. RHAMM regulates the growth and migration of lung adenocarcinoma A549 cell line by regulating Cdc2/CyclinB1 and MMP9 genes[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2150-2163. doi: 10.3934/mbe.2020114

| [1] | R. L. Siegel, K. D. Miller, A. Jemal, Cancer statistics, 2016, CA Cancer J. Clin., 66 (2016), 7-30. |
| [2] | R. Wang, S. Zhang, X. Chen, N. Li, J. Li, R. Jia, et al., EIF4A3-induced circular RNA MMP9 (circMMP9) acts as a sponge of miR-124 and promotes glioblastoma multiforme cell tumorigenesis, Mol. Cancer, 17 (2018), 166. |
| [3] | Y. An, K. L. Furber, S. Ji, Pseudogenes regulate parental gene expression via ceRNA network, J. Cell Mol. Med., 21 (2017), 185-192. |
| [4] |
L. Peng, G. Chen, Z. Zhu, Z. Shen, C. Du, R. Zang, et al., Circular RNA ZNF609 functions as a competitive endogenous RNA to regulate AKT3 expression by sponging miR-150-5p in Hirschsprung's disease, Oncotarget, 8 (2017), 808-818. doi: 10.18632/oncotarget.13656

|
| [5] | Y. Tay, J. Rinn, P. P. Pandolfi, The multilayered complexity of ceRNA crosstalk and competition, Nature, 505 (2014), 344-352. |
| [6] | S. Liu, P. Zhang, Z. Chen, M. Liu, X. Li, H. Tang, MicroRNA-7 downregulates XIAP expression to suppress cell growth and promote apoptosis in cervical cancer cells, FEBS Lett., 587 (2013), 2247-2253. |
| [7] | T. Bian, L. Zheng, D. Jiang, J. Liu, Overexpression of fibronectin type III domain containing 3B is correlated with epithelial-mesenchymal transition and predicts poor prognosis in lung adenocarcinoma, Exp. Ther. Med., 17 (2019), 3317-3326. |
| [8] | M. JD, Molecular pathogenesis of lung cancer, Lung Cancer, 29 (2000), 37-38. |
| [9] | N. Schatz-Siemers, Y. T. Chen, Z. Chen, D. Wang, L. H. Ellenson, Y. N. Du, Expression of the Receptor for Hyaluronic Acid-Mediated Motility (RHAMM) in Endometrial Cancer is Associated With Adverse Histologic Parameters and Tumor Progression, Appl. Immunohistochem. Mol. Morphol., 2019. |
| [10] | C. Tolg, S. R. Hamilton, K. A. Nakrieko, F. Kooshesh, P. Walton, J. B. McCarthy, et al., Rhamm-/- fibroblasts are defective in CD44-mediated ERK1,2 motogenic signaling, leading to defective skin wound repair, J. Cell Biol., 175 (2006), 1017-1028. |
| [11] | E. A. Turley, Purification of a hyaluronate-binding protein fraction that modifies cell social behavior, Biochem. Biophys. Res. Commun., 108 (1982), 1016-1024. |
| [12] | J. P. Lydon, F. J. DeMayo, C. R. Funk, S. K. Mani, A. R. Hughes, C. A. Montgomery, et al., Mice lacking progesterone receptor exhibit pleiotropic reproductive abnormalities, Genes Dev., 9 (1995), 2266-2278. |
| [13] | S. Mohapatra, X. Yang, J. A. Wright, E. A. Turley, A. H. Greenberg, Soluble hyaluronan receptor RHAMM induces mitotic arrest by suppressing Cdc2 and cyclin B1 expression, J. Exp. Med., 183 (1996), 1663-1668. |
| [14] | A. Hamai, H. Benlalam, F. Meslin, M. Hasmim, T. Carré, I. Akalay, et al., Immune surveillance of human cancer: if the cytotoxic T-lymphocytes play the music, does the tumoral system call the tune?, Tissue Antigens, 75 (2010), 1-8. |
| [15] | H. M. Muller, A. Widschwendter, H. Fieg, L. Ivarsson, G. Goebel, E. Perkmann, et al., DNA methylation in serum of breast cancer patients: an independent prognostic marker, Cancer Res., 63 (2003), 7641-7645. |
| [16] | D. Wang, N. Narula, S. Azzopardi, R. S. Smith, A. Nasar, N. K. Altorki, et al., Expression of the receptor for hyaluronic acid mediated motility (RHAMM) is associated with poor prognosis and metastasis in non-small cell lung carcinoma, Oncotarget, 7 (2016), 39957-39969. |
| [17] | F. Augustin, M. Fiegl, T. Schmid, G. Pomme, W. Sterlacci, A. Tzankov, Receptor for hyaluronic acid-mediated motility (RHAMM, CD168) expression is prognostically important in both nodal negative and nodal positive large cell lung cancer, J. Clin. Pathol., 68 (2015), 368-373. |
| [18] | A. Lugli, I. Zlobec, U. Gunthert, P. Minoo, K. Baker, L. Tornillo, et al., Overexpression of the receptor for hyaluronic acid mediated motility is an independent adverse prognostic factor in colorectal cancer, Mod. Pathol., 19 (2006), 1302-1309. |
| [19] | I. Zlobec, L. Terracciano, L. Tornillo, U. Günthert, T. Vuong, J. R. Jass, et al., Role of RHAMM within the hierarchy of well-established prognostic factors in colorectal cancer, Gut, 57 (2008), 1413-1419. |
| [20] | V. H. Koelzer, B. Huber, V. Mele, G. Iezzi, Expression of the hyaluronan-mediated motility receptor RHAMM in tumor budding cells identifies aggressive colorectal cancers, Hum. Pathol., 46 (2015), 1573-1581. |
| [21] | V. Mele, L. Sokol, V. H. Kolzer, D. Pfaff, M. G. Muraro, I. Keller, et al., The hyaluronan-mediated motility receptor RHAMM promotes growth, invasiveness and dissemination of colorectal cancer, Oncotarget, 8 (2017), 70617-70629. |
| [22] | S. T. Buttermore, M. S. Hoffman, A. Kumar, A. Champeaux, S. V. Nicosia, P. A. Kruk, Increased RHAMM expression relates to ovarian cancer progression, J. Ovarian. Res., 10 (2017), 66. |
| [23] | J. M. Song, J. Im, R. S. Nho, Y. H. Han, P. Upadhyaya, F. Kassie, Hyaluronan-CD44/RHAMM interaction-dependent cell proliferation and survival in lung cancer cells, Mol. Carcinog, 58 (2019), 321-333. |
| [24] | K. Kouvidi, A. Berdiaki, M. Tzardi, E. Karousou, A. Passi, D. Nikitovic, et al., Receptor for hyaluronic acid- mediated motility (RHAMM) regulates HT1080 fibrosarcoma cell proliferation via a beta-catenin/c-myc signaling axis, Biochim. Biophys. Acta., 1860 (2016), 814-824. |
| [25] | A. R. Jordan, S. D. Lokeshwar, L. E. Lopez, M. Hennig, J. Chipollini, T. Yates, et al., Antitumor activity of sulfated hyaluronic acid fragments in pre-clinical models of bladder cancer, Oncotarget, 8 (2017), 24262-24274. |
| [26] | S. Matou-Nasri, J. Gaffney, S. Kumar, M. Slevin, Oligosaccharides of hyaluronan induce angiogenesis through distinct CD44 and RHAMM-mediated signalling pathways involving Cdc2 and gamma-adducin, Int. J. Oncol., 35 (2009), 761-773. |
| [27] | P. Pongcharoen, A. Jinawath, R. Tohtong, Silencing of CD44 by siRNA suppressed invasion, migration and adhesion to matrix, but not secretion of MMPs, of cholangiocarcinoma cells, Clin. Exp. Metastasis., 28 (2011), 827-839. |
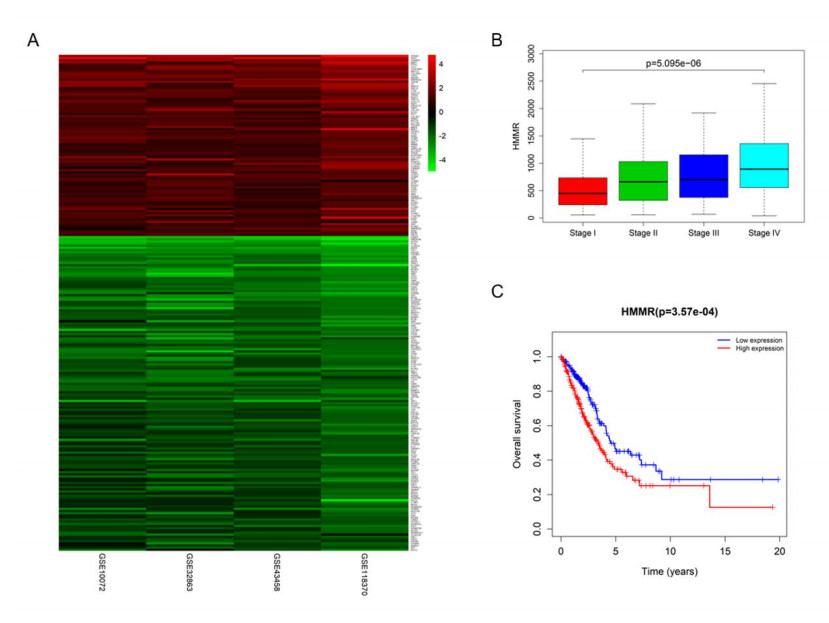
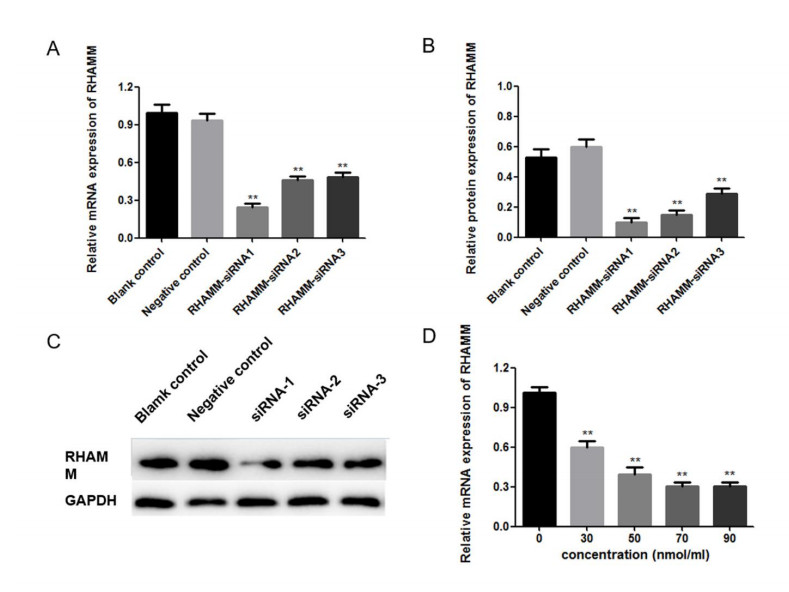
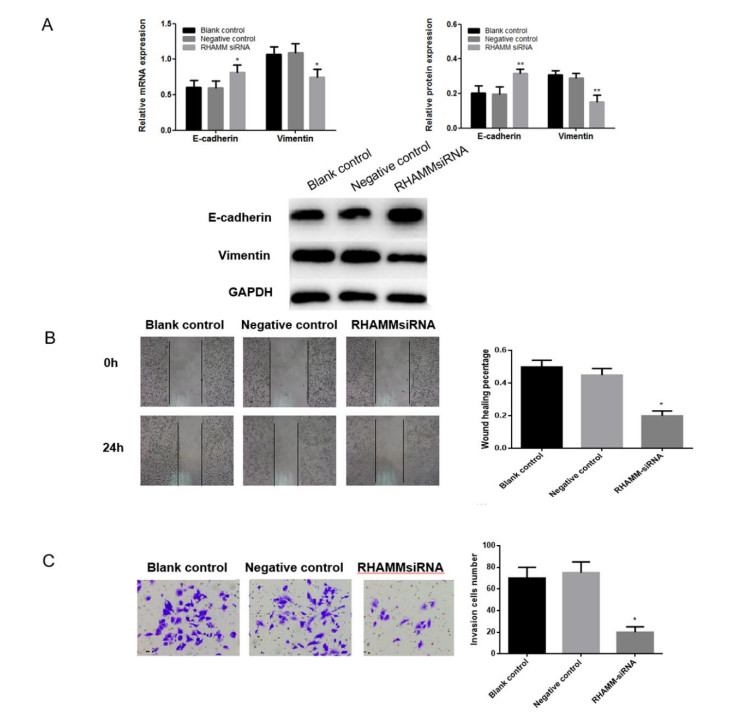
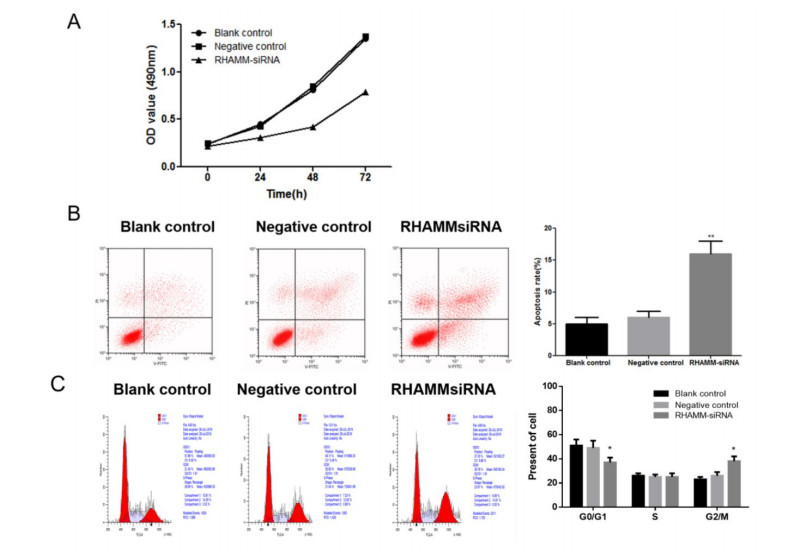
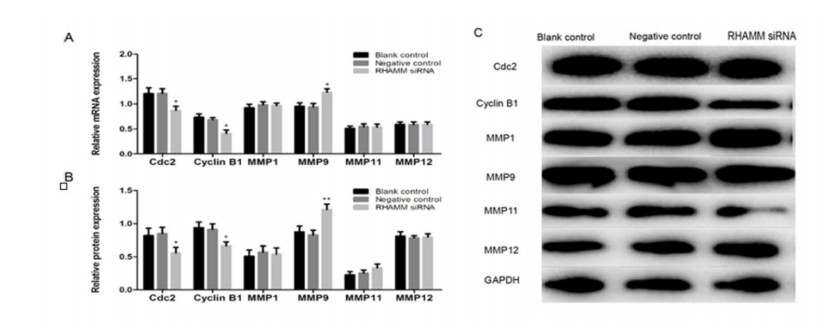
Figures(5) / Tables(1)

Feng Chen, Xuqing Zhu, Jing Zheng, Tingting Xu, Kuan Wu, Chuhui Ru. RHAMM regulates the growth and migration of lung adenocarcinoma A549 cell line by regulating Cdc2/CyclinB1 and MMP9 genes







 DownLoad:
DownLoad: