Citation: Hongying Shu, Wanxiao Xu, Zenghui Hao. Global dynamics of an immunosuppressive infection model with stage structure[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2082-2102. doi: 10.3934/mbe.2020111

| [1] | F. C. Bekkering, C. Stalgis, J. G. McHutchison, J. T. Brouwer, A. S. Perelson, Estimation of early hepatitis C viral clearance in patients receiving daily interferon and ribavirin therapy using a mathematical model, Hepatology, 33 (2001), 419-423. |
| [2] | A. U. Neumann, N. P. Lam, H. Dahari, D. R. Gretch, T. E. Wiley, T. J. Layden, et al., Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-α therapy, Science, 282 (1998), 103-107. |
| [3] | S. A. Whalley, J. M. Murray, D. Brown, G. J. M. Webster, V. C. Emery, G. M. Dusheiko, et al., Kinetics of acute hepatitis B virus infection in humans, J. Exp. Med., 193 (2001), 847-854. |
| [4] | T. W. Chun, L. Stuyver, S. B. Mizell, L. A. Ehler, J. A. Mican, M. Baseler, et al., Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy, Proc. Natl. Acad. Sci. USA, 94 (1997), 13193-13197. |
| [5] | A. S. Perelson, Modelling viral and immune system dynamics, Nat. Rev. Immunol., 2 (2002), 28-36. |
| [6] | N. L. Komarova, E. Barnes,P. Klenerman, D. Wodarz, Boosting immunity by antiviral drug therapy: a simple relationship among timing, efficacy, and success, Proc. Natl. Acad. Sci. USA, 100 (2003), 1855-1860. |
| [7] | E. S. Rosenberg, M. Altfeld, S. H. Poon, M. N. Phillips, B. M. Wilkes, R. L. Eldridge, et al., Immune control of HIV-1 after early treatment of acute infection, Nature, 407 (2000), 523-526. |
| [8] | A. Fenton, J. Lello, M. B. Bonsall, Pathogen responses to host immunity: the impact of time delays and memory on the evolution of virulence, Proc. R. Soc. B, 273 (2006), 2083-2090. |
| [9] | G. M. Ortiz, J. Hu, J. A. Goldwitz, R. Chandwani, M. Larsson, N. Bhardwaj, et al., Residual viral replication during antiretroviral therapy boosts human immunodeficiency virus type 1-specific CD8+ T-cell responses in subjects treated early after infection, J. Virol., 76 (2002), 411-415. |
| [10] | H. Shu, L. Wang, J. Watmough, Sustained and transient oscillations and chaos induced by delayed antiviral immune response in an immunosuppressive infection model, J. Math. Biol., 68 (2014), 477-503. |
| [11] | J. K. Hale, S. M. V. Lunel, Introduction to Functional Differential Equations, Springer-Verlag, New York, 1993. |
| [12] | X. Pan, Y. Chen, H. Shu, Rich dynamics in a delayed HTLV-I infection model: stability switch, multiple stable cycles, and torus, J. Math. Anal. Appl., 479 (2019), 2214-2235. |
| [13] | H. Shu, X. Hu, L. Wang, J. Watmough, Delay induced stability switch, multitype bistability and chaos in an intraguild predation model, J. Math. Biol., 71 (2015), 1269-1298. |
| [14] | J. K. Hale, Asymptotic Behavior of Dissipative Systems, AMS, Providence, 1988. |
| [15] | H. Thieme, Uniform persistence and permanence for non-autonomous semiflows in population biology, Math. Biosci., 166 (2000), 173-201. |
| [16] | H. Shu, L. Wang, J. Watmough, Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL immune responses, SIAM J. Appl. Math., 73 (2013), 1280-1302. |
| [17] | E. Beretta, Y. Kuang, Geometric stability switch criteria in delay differential systems with delay dependent parameters, SIAM J. Math. Anal., 33 (2002), 1144-1165. |
| [18] | B. D. Hassard, N. D. Kazarinoff, Y. H. Wan, Theory and Applications of Hopf Bifurcation, Cambridge University Press, Cambridge, 1981. |
| [19] | J. Wu, Symmetric functional differential equations and neural networks with memory, Trans. Amer. Math. Soc., 350 (1998), 4799-4838. |
| [20] | K. Engelborghs, T. Luzyanina, G. Samaey, DDE-BIFTOOL v. 2.00: A Matlab package for bifurcation analysis of delay differential equations, Technical Report TW-330, K. U. Leuven, Belgium, 2001. |
| [21] | K. Engelborghs, T. Luzyanina, D. Roose, Numerical bifurcation analysis of delay differential equations using DDE-BIFTOOL, ACM Trans. Math. Software, 28 (2002), 1-21. |
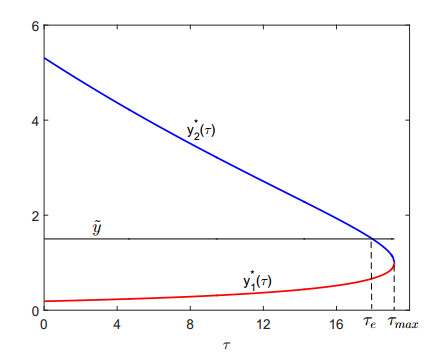
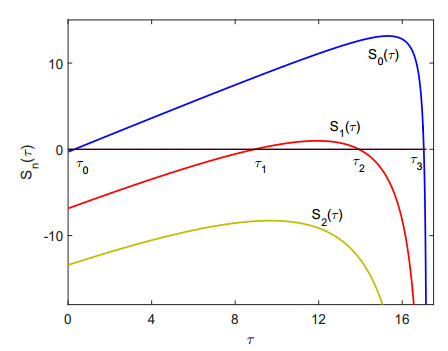
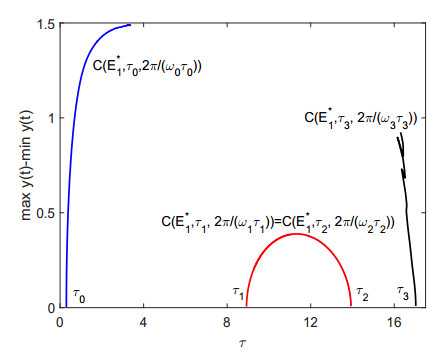
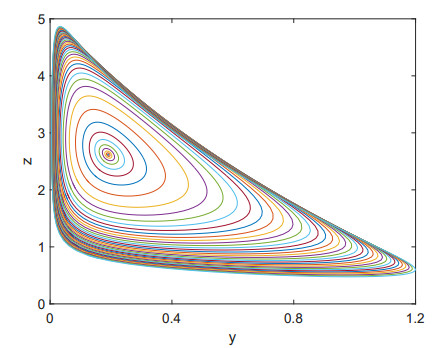
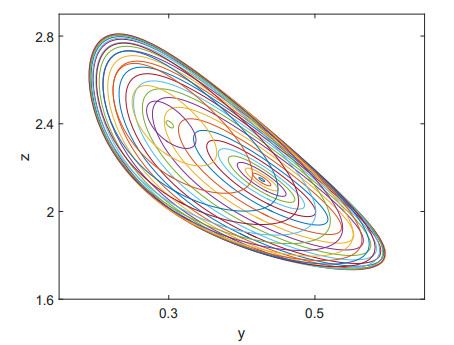
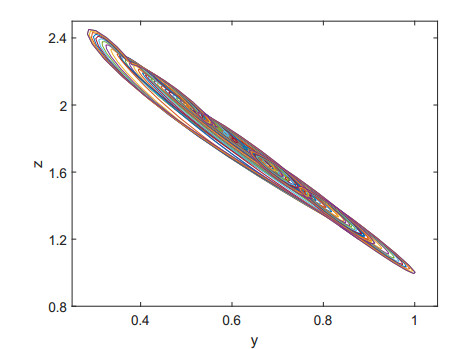

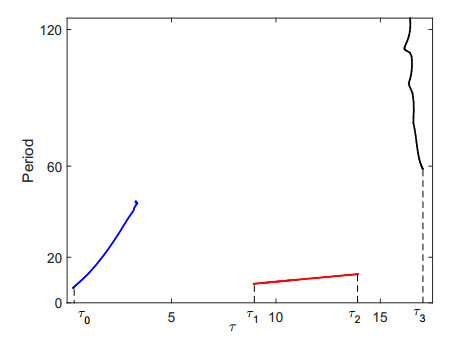
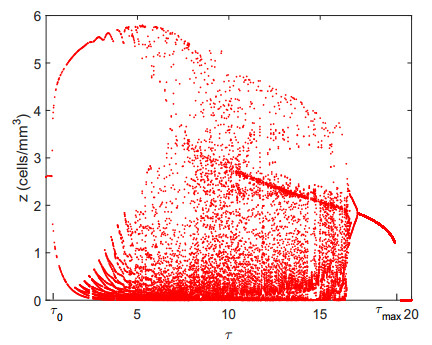
Figures(9)

Hongying Shu, Wanxiao Xu, Zenghui Hao. Global dynamics of an immunosuppressive infection model with stage structure[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2082-2102. doi: 10.3934/mbe.2020111







 DownLoad:
DownLoad: