Citation: Alejandro Rincón, Fabiola Angulo, Fredy E. Hoyos. Analysis of a generalized Fujikawa’s growth model[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2103-2137. doi: 10.3934/mbe.2020112

| [1] | J. Baranyi, T.A. Roberts, P. McClure, A non-autonomous differential equation to model bacterial growth, Int. J. Food Microbiol., 10 (1993), 43-59. |
| [2] | M.H. Zwietering, I. Jongenburger, F.M Rombouts, K. Van't Riet, Modeling of the bacterial growth curve, Appl. Environ. Microb., 56 (1990), 1875-1881. |
| [3] | S. Perni, P.W. Andrew, G. Shama, Estimating the maximum growth rate from microbial growth curves: Definition is everything, Int. J. Food Microbiol., 22 (2005), 491-495. |
| [4] | S. Basak, P. Guha, Modelling the effect of essential oil of betel leaf (Piper betle L.) on germination, growth, and apparent lag time of Penicillium expansum on semi-synthetic media, Int. J. Food Microbiol., 215 (2015), 171-178. |
| [5] | P.R. Santos, I.C. Tessaro, L.D. Ferreira, Integrating a kinetic microbial model with a heat transfer model to predict Byssochlamys fulva growth in refrigerated papaya pulp, J. Food Eng., 118 (2013), 279-288. |
| [6] | P.R. Santos, I.C. Tessaro, L.D. Ferreira, Modeling and simulation of Byssochlamys fulva growth on papaya pulp subjected to evaporative cooling, Chem. Eng. Sci., 114 (2014), 134-143. |
| [7] | R.M. de Castro, J.R. de Souza, A.F da Eira, Digital monitoring of mycelium growth kinetics and vigor of shiitake (Lentinula edodes (Berk.) Pegler) on agar medium, Braz. J. Microbiol., 37 (2006), 90-95. |
| [8] | A. Talkington, R. Durrett, Estimating tumor growth rates in vivo, Bull. Math. Biol., 77 (2015), 1934-1954. |
| [9] | C.P. Birch, A new generalized logistic sigmoid growth equation compared with the Richards growth equation, Ann. Bot. London, 83 (1999), 713-723. |
| [10] | X. Yin, J. Goudriaan, E.A. Lantinga, J. Vos, H.J. Spiertz, A flexible sigmoid function of determinate growth, Ann. Bot. London, 91 (2003), 361-37. |
| [11] | J. Ukalska, S. Jastrzebowski, Sigmoid growth curves, a new approach to study the dynamics of the epicotyl emergence of oak, Folia For. Pol., Ser. A, 61 (2019), 30-41. |
| [12] | J.G. Garca, R. Ramrez, R. Nez, J.A. Hidalgo, Dataset on growth curves of Boer goats fitted by ten non-linear functions, Data Brief, 23 (2019), 1-10. |
| [13] | J.M. Coyne, K. Matilainen, D.P. Berry, M.L. Sevn, E.A. Mantysaari, J.Juga, et al., Estimation of genetic (co)variances of Gompertz growth function parameters in pigs, J. Anim. Breed Genet., 134 (2017), 136-143. |
| [14] | R.C. Bruce, Application of the Gompertz function in studies of growth in dusky salamanders (Plethodontidae: Desmognathus), Copeia, 104 (2016), 94-100. |
| [15] | A. Silveira, J.R. Moreno, M.J. Correia, V. Ferro, A method for the rapid evaluation of leather biodegradability during the production phase, Waste Manage., 87 (2019), 661-661. |
| [16] | H. Fujikawa, A. Kai, S. Morozumi, A new logistic model for Escherichia coli growth at constant and dynamic temperatures, Int. J. Food Microbiol., 21 (2004), 501-509. |
| [17] | P. Vadasz, A.S. Vadasz, Predictive modeling of microorganisms: LAG and LIP in monotonic growth, Int. J. Food Microbiol., 102 (2005), 257-257. |
| [18] | S. Brown, An estimate of the duration of the lag phase of the logistic growth curve, Ann. West Univ. Timisoara, 17 (2014), 25-32. |
| [19] | A. Tsoularis, J. Wallace, Analysis of logistic growth models, Math. Biosci., 179 (2002), 21-55. |
| [20] | S. Ohnishi, T. Yamakawa, T. Akamine, On the analytical solution for the Putter-Bertalanffy growth equation, J. Theor. Biol., 343 (2014), 174-177. |
| [21] | P. Vadasz, A.S. Vadasz, The neoclassical theory of population dynamics in spatially homogeneous environments. (I) Derivation of universal laws and monotonic growth, Physica A, 309 (2002), 329-359. |
| [22] | F. Poschet, K.M. Vereecken, A.H. Geeraerd, B.M. Nicolai, J.F. Van Impe, Analysis of a novel class of predictive microbial growth models and application to coculture growth, Int. J. Food Microbiol., 100 (2005), 107-124. |
| [23] | J.F. Van Impe, F. Poschet, A.H. Geeraerd, K.M. Vereecken, Towards a novel class of predictive microbial growth models, Int. J. Food Microbiol., 100 (2005), 97-105. |
| [24] | I.A. Swinnen, K. Bernaerts, E.J. Dens, A.H. Geeraerd, J.F. Van Impe, Predictive modelling of the microbial lag phase: A review, Int. J. Food Microbiol., 94 (2004), 137-159. |
| [25] | A. Di Crescenzo, S. Spina, Analysis of a growth model inspired by Gompertz and Korf laws, and an analogous birth-death process, Math. Biosci, 282 (2016), 121-134. |
| [26] | I. Mytilinaios, M. Salih, H.K. Schofield, R.J. Lambert, Growth curve prediction from optical density data, Int. J. Food Microbiol., 154 (2012), 169-176. |
| [27] | B.M. Nicolai, J.F. Van Impe, B. Verlinden, T. Martens, J. Vandewalle, J. De Baerdemaeker, Predictive modelling of surface growth of lactic acid bacteria in vacuum-packed meat, Int. J. Food Microbiol., 10 (1993), 229-238. |
| [28] | H. Fujikawa, S. Morozumi, Modeling Staphylococcus aureus growth and enterotoxin production in milk, Int. J. Food Microbiol., 23 (2006), 260-267. |
| [29] | J. Baranyi, T.A. Roberts, Mathematics of predictive food Microbiol.ogy, Int. J. Food Microbiol., 26 (1995), 199-218. |
| [30] | P.R Koya, A.T. Goshu, Generalized mathematical model for biological growths, Open J. Modell. Simul., 1 (2013), 42-53. |
| [31] | J. Baranyi, T.A. Roberts, A dynamic approach to predicting bacterial growth in food, Int. J. Food Microbiol., 23 (1994), 277-294. |
| [32] | F. Baty, M. L. Delignette-Muller, Estimating the bacterial lag time: Which model, which precision?, Int. J. Food Microbiol., 91 (2004), 261-277. |
| [33] | A.W. Mayo, M. Muraza, J. Norbert, Modelling nitrogen transformation and removal in mara river basin wetlands upstream of lake Victoria, Phys. Chem. Earth, 105 (2018), 136-146. |
| [34] | K. Dutta, V.V. Dasu, B. Mahanty, A.A. Prabhu, Substrate inhibition growth kinetics for cutinase producing pseudomonas cepacia using tomato-peel extracted cutin, Chem. Biochem. Eng. Q, 29 (2015), 437-445. |
| [35] | M. Li, Y. Li, X. Huang, G. Zhao, W. Tian, Evaluating growth models of Pseudomonas spp. in seasoned prepared chicken stored at different temperatures by the principal component analysis (PCA), Int. J. Food Microbiol., 40 (2014), 41-47. |
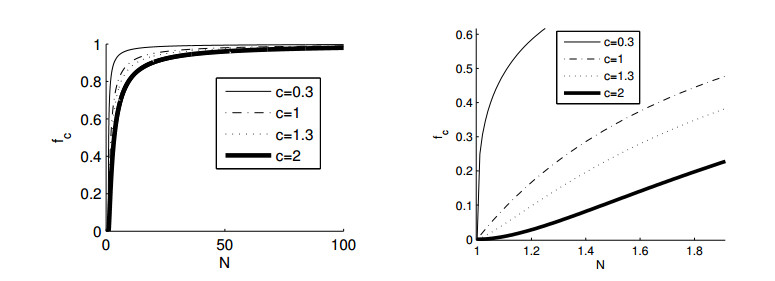
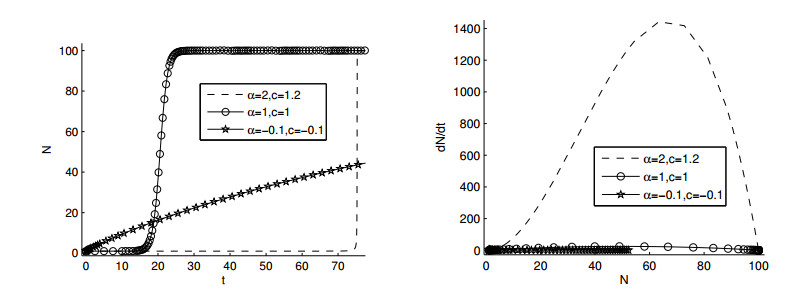
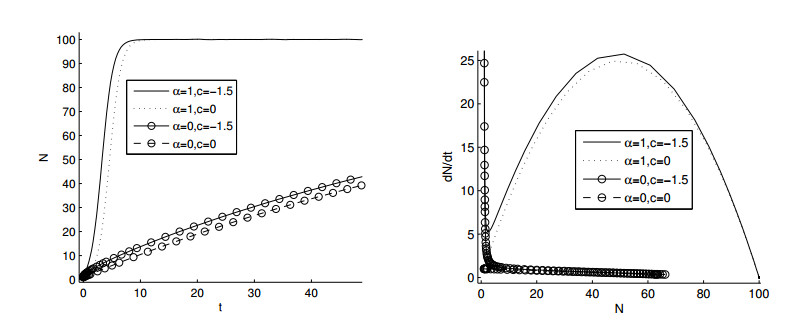
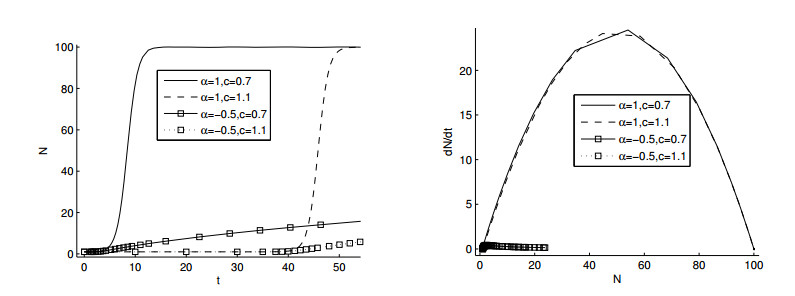
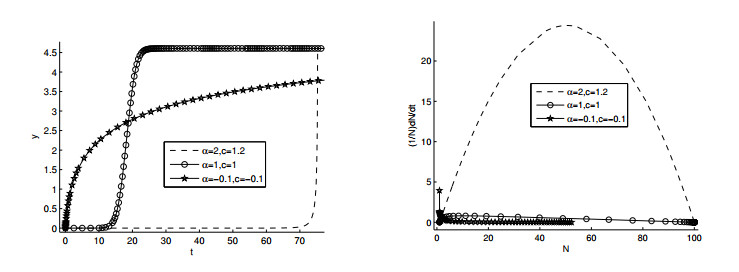
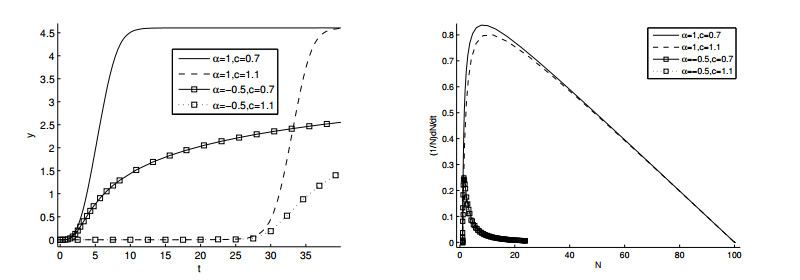
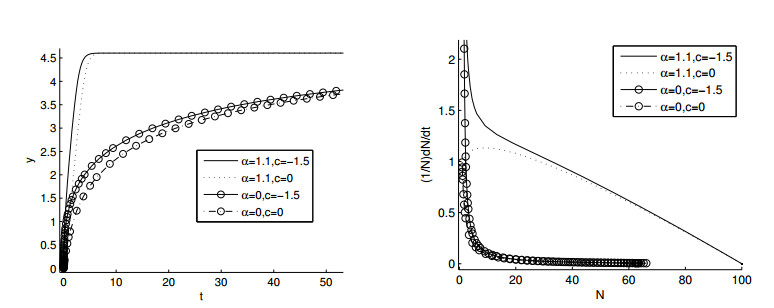
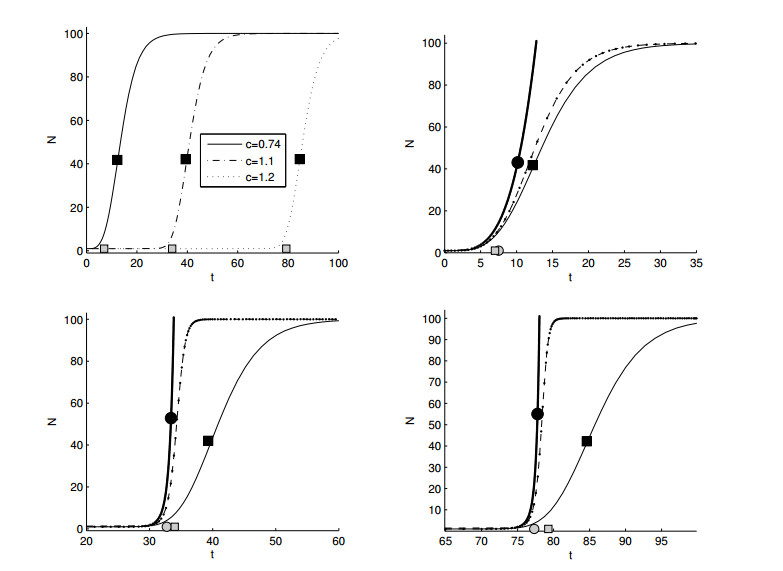
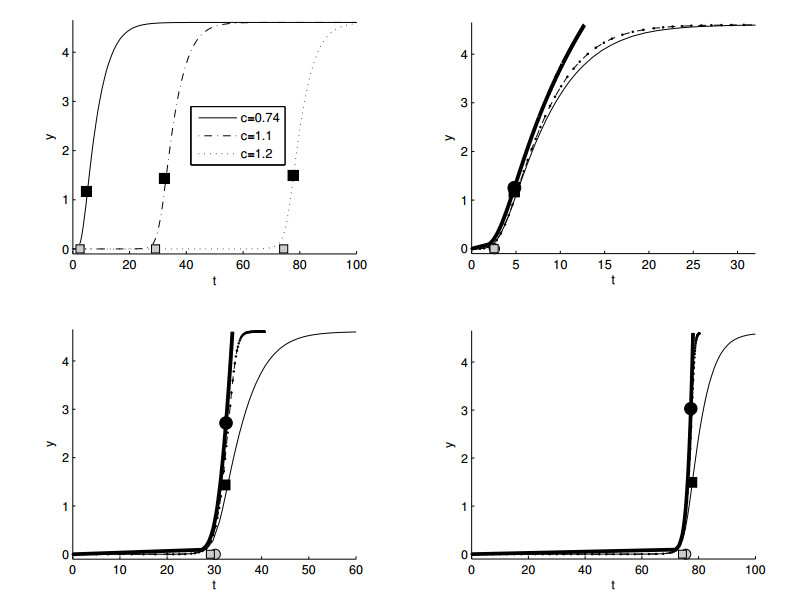
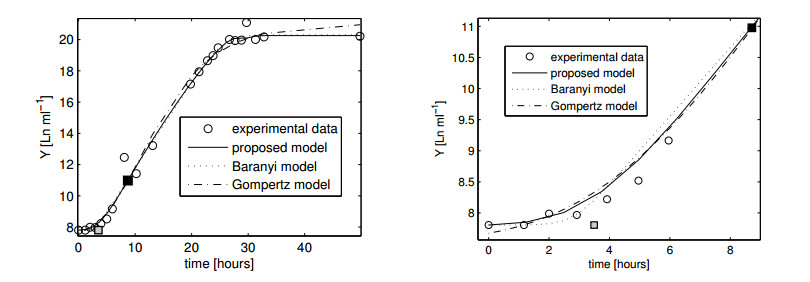
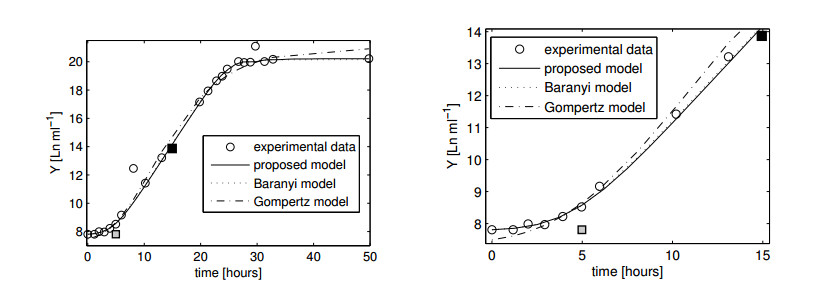
Figures(11)

Alejandro Rincón, Fabiola Angulo, Fredy E. Hoyos. Analysis of a generalized Fujikawa’s growth model[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2103-2137. doi: 10.3934/mbe.2020112







 DownLoad:
DownLoad: