Citation: Chang-Yuan Cheng, Shyan-Shiou Chen, Xingfu Zou. On an age structured population model with density-dependent dispersals between two patches[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4976-4998. doi: 10.3934/mbe.2019251

| [1] | A. E. Abdllaoui, P. Auger, B. W. Kooi, et al., Effects of density-dependent migrations on stability of a two-patch predator-prey model, Math. Biosci., 210 (2007), 335–354. |
| [2] | S. A. Levin, Dispersion and population interactions, Amer. Natur., 108 (197), 207–228. |
| [3] | J. A. J. Metz and O. Diekmann, The Dynamics of Physiologically Structured Populations, Springer-Verlag, New York, 1986. |
| [4] | J. W. H. So, J. Wu and X. Zou, Structured population on two patches: modeling dispersal and delay, J. Math. Biol., 43 (2001), 37–51. |
| [5] | P. Weng, C. Xiao and X. Zou, Rich dynamics in a non-local population model over three patches, Nonlinear Dynam., 59 (2010), 161–172. |
| [6] | D. Xu, Global dynamics and Hopf bifurcation of a structured population model, Nonl. Anal. Real World Appl., 6 (2005), 461–476. |
| [7] | C. Yu, J. Wei and X. Zou, Bifurcation analysis in an age-structured model of a single species living in two identical patches, Appl. Math. Model., 34 (2010), 1068–1077. |
| [8] | A. J. Terry, Dynamics of a structured population on two patches, J. Math. Anal. Appl., 378 (2011), 1–15. |
| [9] | R. Cressman and V. Křivan, Two-patch population models with adaptive dispersal: the effects of varying dispersal speed, J. Math. Biol., 67 (2013), 329–358. |
| [10] | W. Wang and Y. Tacheuchi, Adaption of prey and predators between patches, J. Theo. Biol., 258 (2011), 603–613. |
| [11] | X. Zhang and W. Wang, Importance of dispersal adaption of two competitive populations between patches, Ecol. Model., 222 (2011), 11–20. |
| [12] | C.Huang, Z.Yang, T.Yi, et al., On the basins of attraction for a class of delay differential equations with non-monotone bistable nonlinearities, J. Diff. Eq., 256 (2014), 2101–2114. |
| [13] | G. Rost and J. Wu, Domain-decomposition method for the global dynamics of delay differential equations with unimodal feedback, Proc. R. Soc. A, 463 (2007), 2655–2665. |
| [14] | Y. Yuan and X. Q. Zhao, Global stability for non-monotone delay equations (with application to a model of blood cell production), J. Diff. Eq., 252 (2012), 2189–2209. |
| [15] | A. J. Terry, Impulsive culling of a structured population on two patches, J. Math. Biol., 61 (2010), 843–875. |
| [16] | H. C. J. Godfray, L. Partridge and P. H. Harvey, Clutch size, Annu. Rev. Ecol., 22 (1991), 409–429. |
| [17] | B. K. Sandercock, Incubation capacity and clutch size determination in two calidrine sandpipers: a test of the four-egg threshold, Oecologia, 110 (1997), 50–59. |
| [18] | B. A. Shanbhag, Reproductive strategies in the lizard, Calotes versicolor, Curr. Sci., 84 (2003), 646–652. |
| [19] | J. K. Hale and S. M. Verduyn Lunel, Introduction to functional differential equations, Springer, New York, 1993. |
| [20] | H. L. Smith, Monotone Dynamical Systems: An Introduction to the theory of Competitive and Cooperative Systems, Mathematical Surveys and Monographs, vol. 41, AMS, Providence, RI, 1995. |
| [21] | H. R. Thieme, Persistence under relaxed point-dissipativity (with application to an endemic model), SIAM J. Math. Anal., 24 (1993), 407–435. |
| [22] | M. A. McPeek and R. D. Holt, The evolution of dispersal in spatially and temporally varying environments, Amer. Natur., 140 (1992), 1010–1027. |
| [23] | J. Dockery, V. Hutson, K. Mischaikow, et al., The evolution of slow dispersal rates: a reaction-diffusion model, J. Math. Biol., 37 (1998), 61–83. |
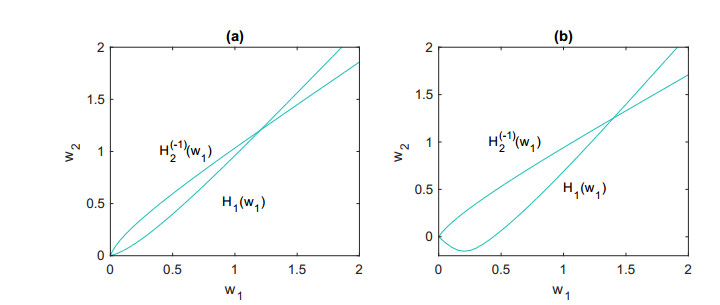
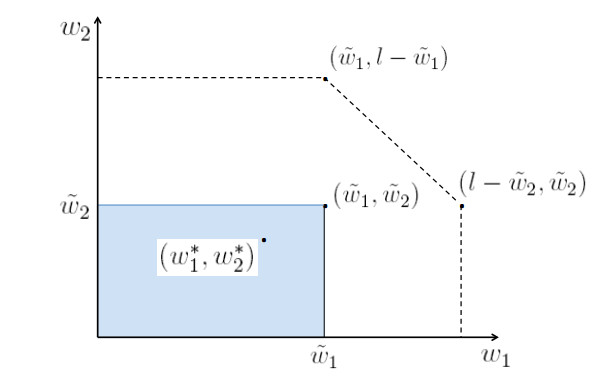
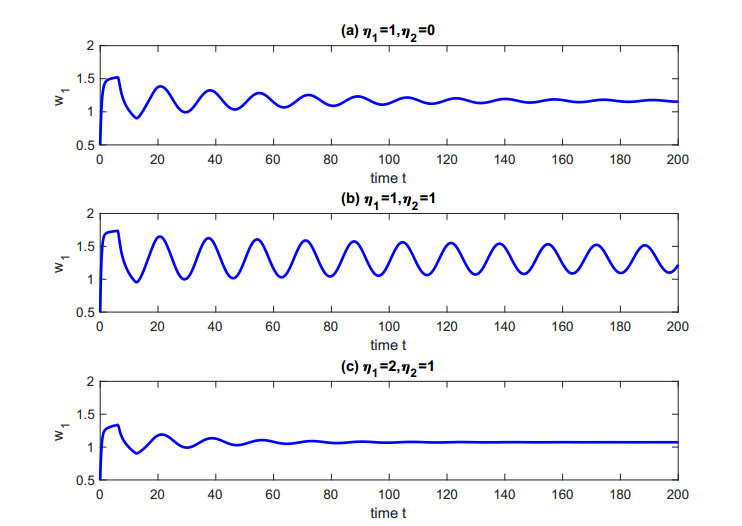
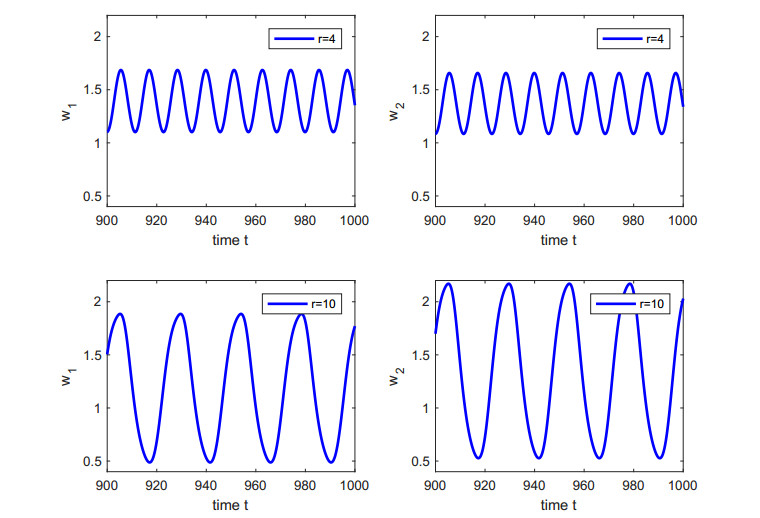
Figures(5)

Chang-Yuan Cheng, Shyan-Shiou Chen, Xingfu Zou. On an age structured population model with density-dependent dispersals between two patches[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4976-4998. doi: 10.3934/mbe.2019251







 DownLoad:
DownLoad: