Citation: Mondal Hasan Zahid, Christopher M. Kribs. Ebola: Impact of hospital's admission policy in an overwhelmed scenario[J]. Mathematical Biosciences and Engineering, 2018, 15(6): 1387-1399. doi: 10.3934/mbe.2018063

| [1] | [ Central Intelligence Agency, CIA. The World Factbook, Update date: 03-01-2016, https://www.cia.gov/library/publications/the-world-factbook/fields/2227.html, Access date: 03-07-2017. |
| [2] | [ M. D. Ahmad, M. Usman, A. Khan and M. Imran, Control analysis of Ebola disease with control strategies of quarantine and vaccination, Infectious Disease of Poverty, 5 (2016), 72. |
| [3] | [ M. Ajelli, S. Parlamento1, D. Bome, A. Kebbi, A. Atzori, C. Frasson, G. Putoto, D. Carraro and S. Merler, The 2014 Ebola virus disease outbreak in Pujehun, Sierra Leone: epidemiology and impact of interventions, BMC Medicine, 13 (2015), 281. |
| [4] | [ C. L. Althaus, Estimating the reproduction number of Ebola Virus (EBOV) during the 2014 outbreak in West Africa, PLOS Currents Outbreaks, 2014. |
| [5] | [ R. Ansumana, K. H. Jacobsen, M. Idris, H. Bangura, M. Boie-Jalloh, J. M. Lamin, S. Sesay and F. Sahr, Ebola in Freetown Area, Sierra Leone A case study of 581 patients, New England Journal of Medicine, 372 (2015), 587-588. |
| [6] | [ A. G. Buseh, P. E. Stevens, M. Bromberg and S. T. Kelber, The Ebola epidemic in West Africa: Challenges, opportunities, and policy priority areas, Nursing Outlook, 63 (2015), 30–40. http://dx.doi.org/10.1016/j.outlook.2014.12.013. |
| [7] | [ G. Chowell, N. W. Hengartner, C. Castillo-Chavez, P. W. Fenimore and J. M. Hyman, The basic reproductive number of Ebola and the effects of public health measures: the cases of Congo and Uganda, Journal of Theoretical Biology, 229 (2004), 119-126. |
| [8] | [ O. Diekmann, J. A. P. Heesterbeek and J. A. J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol, 28 (1990), 365-382. |
| [9] | [ J. M. Drake, R. B. Kaul, L. W. Alexander, S. M. Oegan, A. M. Kramer, J. T. Pulliam, M. J. Ferrari and A. W. Park, Ebola cases and health system demand in liberia, PLoS Biology, 13 (2015), e1002056. |
| [10] | [ T. W. Geisbert, L. E. Hensley, P. B. Jahrling, T. Larsen, J. B. Geisbert, J. Paragas, H. A. Young, T. M. Fredeking, W. E. Rote and G. P. Vlasuk, Treatment of Ebola virus infection with a recombinant inhibitor of factor VIIa/tissue factor: a study in rhesus monkeys, The Lancet, 362 (2003), 1953-1958. |
| [11] | [ Ministry of Health, Uganda, Uganda Hospital and Health Centre IV Census Survey, 2014,198. |
| [12] | [ P. Kazanjian, Ebola in antiquity, Clinical Infectious Disease, 61 (2015), 963-968. |
| [13] | [ Dr. A. I. Khan, Chief Physician and Head, Hospitals, icddr, b, Dhaka, Bangladesh, Personal communication, October 11, 2017. |
| [14] | [ Medecins Sans Frontieres, International Response to West Africa Ebola Epidemic Dangerously Inadequate, August 15, 2014. https://www.msf.org/international-response-west-africa-ebola-epidemic-dangerously-inadequate, Access date: 2018-06-01. |
| [15] | [ Sylvie Diane Djiomba Njankou and Farai Nyabadza, Modelling the potential impact of limited hospital beds on Ebola virus disease dynamics, Mathematical Methods in the Applied Sciences, (2018), 1-17. |
| [16] | [ World Health Organization, Ebola Situation Report, June 10, 2016. http://who.int/csr/disease/ebola/en/. |
| [17] | [ World Health Organization, Health Statistics and Information Systems, Update date: 07-01-2016, http://www.who.int/healthinfo/global_burden_disease/metrics_daly/en/, Access date: 03-03-2017. |
| [18] | [ World Health Organization, Liberia: Ebola Treatment Centre Sets A New Pace, October 2014. http://www.who.int/features/2014/liberia-ebola-island-clinic/en/, Access date 2018-06-01. |
| [19] | [ World Health Organization, Why the Ebola outbreak has been underestimated, August 22, 2014. http://www.who.int/mediacentre/news/ebola/22-august-2014/en/, Access date 2018-06-01. |
| [20] | [ E. Qin, J. Bi, M. Zhao, Y. Wang, T. Guo, T. Yan, Z. Li, J. Sun, J. Zhang, S. Chen, Y. Wu, J. Li and Y. Zhong, Clinical features of patients with Ebola virus disease in Sierra Leone, Clinical Infectious Diseases, 61 (2015), 491-495. |
| [21] | [ J. A. Salomon, J. A. Haagsma, A. Davis, C. M. de Noordhout, S. Polinder, A. H. Havelaar, A. Cassini, B. Devleesschauwer, M. Kretzschmar, N. Speybroeck and C. J. L. Murray, Disability weights for the Global Burden of Disease 2013 study, The Lancet, 3 (2015), 712-723. |
| [22] | [ J. S. Schieffelin, J. G. Shaffer, A. Goba, M. Gbakie, S. K. Gire, A. Colubri, R. S. G. Sealfon, L. Kanneh, A. Moigboi, M. Momoh, M. Fullah, L. M. Moses, B. L. Brown, K. G. Andersen, S. Winnicki, S. F. Schaffner, D. J. Park, N. L. Yozwiak, P.-P. Jiang, D. Kargbo, S. Jalloh, M. Fonnie, V. Sinnah, I. French, A. Kovoma, F. K. Kamara, V. Tucker, E. Konuwa, J. Sellu, I. Mustapha, M. Foday, M. Yillah, F. Kanneh, S. Saffa, J. L. B. Massally, M. L. Boisen, L. M. Branco, M. A. Vandi, D. S. Grant, C. Happi, S. M. Gevao, T. E. Fletcher, R. A. Fowler, D. G. Bausch, P. C. Sabeti, S. H. Khan and R. F. Garry, Clinical illness and outcomes in patients with Ebola in Sierra Leone, New England Journal of Medicine, 371 (2014), 2092-2100. |
| [23] | [ National Center for Health Statistics, Health, United States, 2015: With Special Feature on Racial and Ethnic Health Disparities, Hyattsville, MD. 2016. |

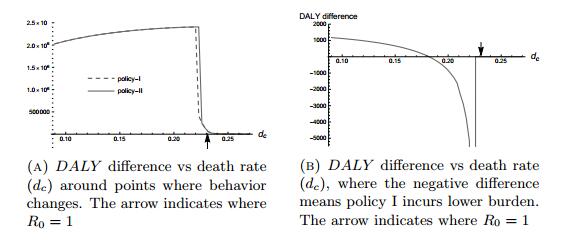
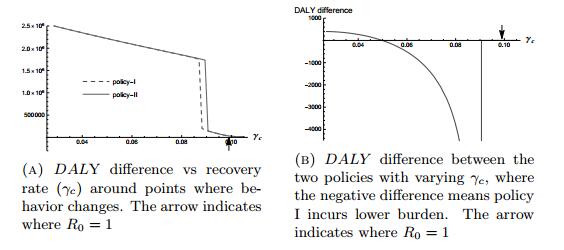
Figures(7) / Tables(3)

Mondal Hasan Zahid, Christopher M. Kribs. Ebola: Impact of hospital's admission policy in an overwhelmed scenario[J]. Mathematical Biosciences and Engineering, 2018, 15(6): 1387-1399. doi: 10.3934/mbe.2018063







 DownLoad:
DownLoad: