Citation: Carole Guillevin, Rémy Guillevin, Alain Miranville, Angélique Perrillat-Mercerot. Analysis of a mathematical model for brain lactate kinetics[J]. Mathematical Biosciences and Engineering, 2018, 15(5): 1225-1242. doi: 10.3934/mbe.2018056

| [1] | [ A. Aubert,R. Costalat, Interaction between astrocytes and neurons studied using a mathematical model of compartmentalized energy metabolism, Journal of Cerebral Blood Flow & Metabolism, 25 (2005): 1476-1490. |
| [2] | [ A. Aubert,R. Costalat,P. Magistretti,J. Pierre,L. Pellerin, Brain lactate kinetics: modeling evidence for neuronal lactate uptake upon activation, Proceedings of the National Academy of Sciences of the United States of America, 102 (2005): 16448-16453. |
| [3] | [ M. Cloutier,F. B. Bolger,J. P. Lowry,P. Wellstead, An integrative dynamic model of brain energy metabolism using in vivo neurochemical measurements, Journal of Computational Neuroscience, 27 (2009): 391-414. |
| [4] | [ R. Costalat,J.-P. Françoise,C. Menuel,M. Lahutte,J.-N. Vallée,G. De Marco,J. Chiras,R. Guillevin, Mathematical modeling of metabolism and hemodynamics, Acta Biotheoretica, 60 (2012): 99-107. |
| [5] | [ C. E. Griguer,C. R. Oliva,G. Y. Gillespie, Glucose metabolism heterogeneity in human and mouse malignant glioma cell lines, Journal of Neuro-oncology, 74 (2005): 123-133. |
| [6] | [ R. Guillevin,C. Menuel,J.-N. Vallée,J.-P. Françoise,L. Capelle,C. Habas,G. De Marco,J. Chiras,R. Costalat, Mathematical modeling of energy metabolism and hemodynamics of WHO grade Ⅱ gliomas using in vivo MR data, Comptes rendus biologies, 334 (2011): 31-38. |
| [7] | [ M. Lahutte-Auboin, R. Costalat, J.-P. Françoise, R. Guillevin, Dip and Buffering in a fast-slow system associated to Brain Lactacte Kinetics, preprint, arXiv: 1308.0486. |
| [8] | [ M. Lahutte-Auboin,R. Guillevin,J.-P. Françoise,J.-N. Vallée,R. Costalat, On a minimal model for hemodynamics and metabolism of lactate : application to low grade glioma and therapeutic strategies, Acta Biotheoretica, 61 (2013): 79-89. |
| [9] | [ P. J. Magistretti,I. Allaman, A cellular perspective on brain energy metabolism and functional imaging, Neuron, 86 (2015): 883-901. |
| [10] | [ S. Mangia,G. Garreffa,M. Bianciardi,F. Giove,F. Di Salle,B. Maraviglia, The aerobic brain: Lactate decrease at the onset of neural activity, Neuroscience, 118 (2003): 7-10. |
| [11] | [ J. R. Mangiardi,P. Yodice, Metabolism of the malignant astrocytoma, Neurosurgery, 26 (1990): 1-19. |
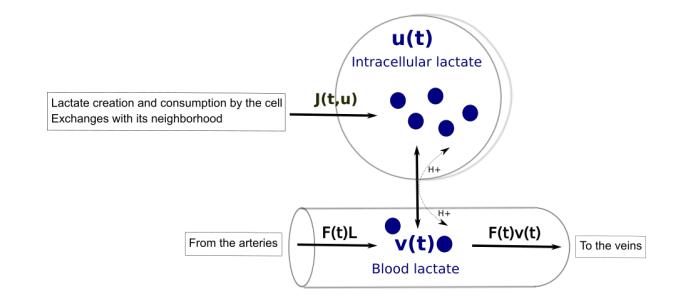
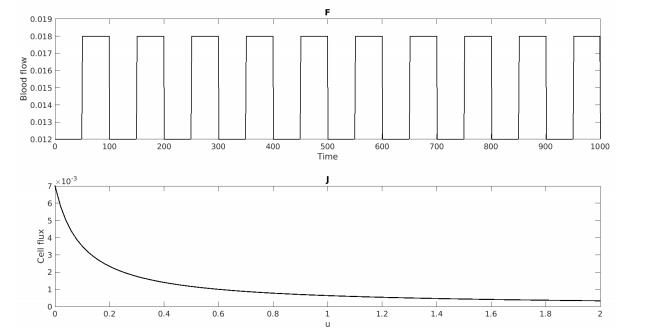
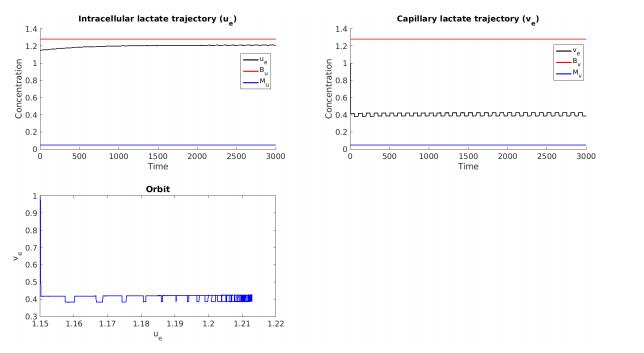
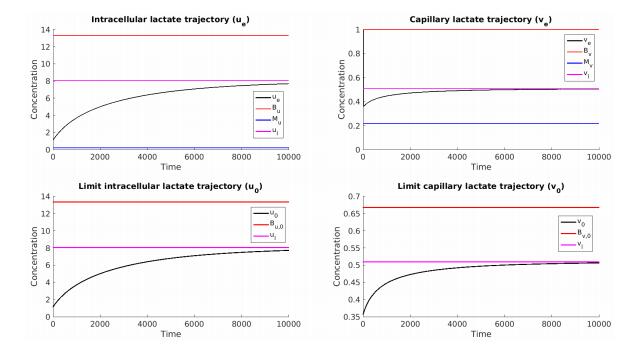
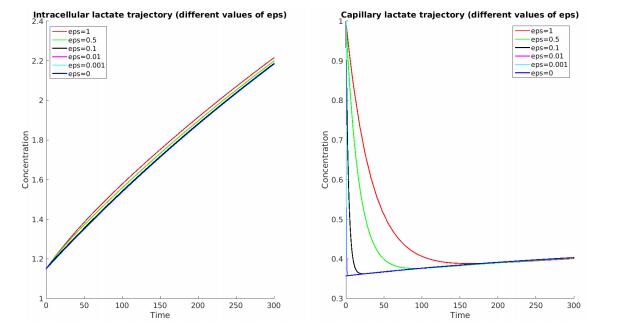
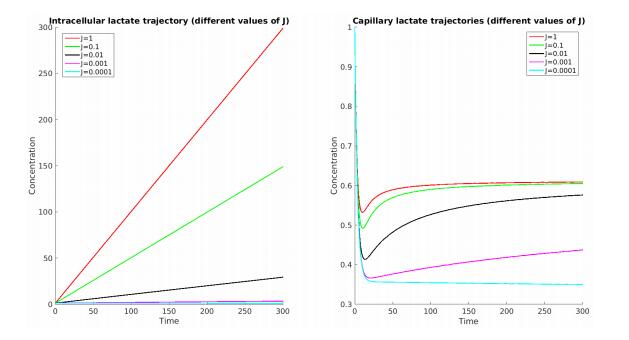
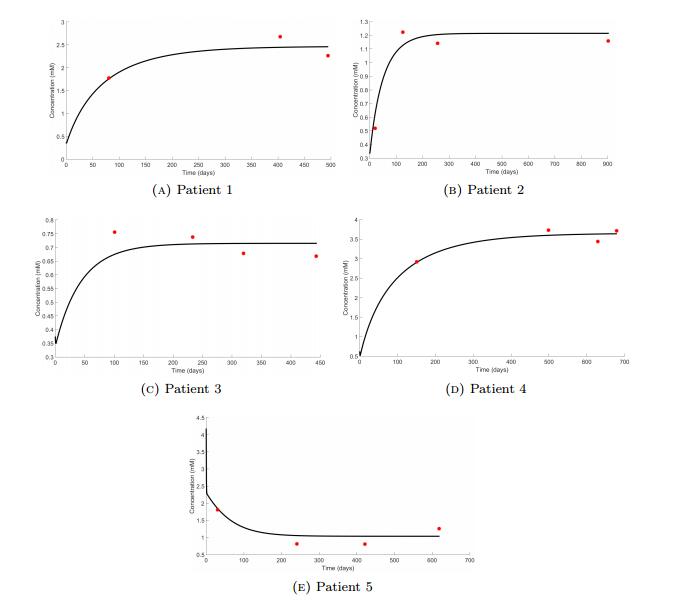
Figures(7) / Tables(5)

Carole Guillevin, Rémy Guillevin, Alain Miranville, Angélique Perrillat-Mercerot. Analysis of a mathematical model for brain lactate kinetics[J]. Mathematical Biosciences and Engineering, 2018, 15(5): 1225-1242. doi: 10.3934/mbe.2018056







 DownLoad:
DownLoad: