Citation: OPhir Nave, Shlomo Hareli, Miriam Elbaz, Itzhak Hayim Iluz, Svetlana Bunimovich-Mendrazitsky. BCG and IL − 2 model for bladder cancer treatment with fast and slow dynamics based on SPVF method—stability analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5346-5379. doi: 10.3934/mbe.2019267

| [1] | V. M. Gol'dshtein and V. A. Sobolev, Singularity theory and some problems of functional analysis, Amer. Math. Soc., 1 (1992), 73–92. |
| [2] | V. I. Babushok and V. M. Gol'dshtein, Structure of the thermal explosion limit, Combust. Flame, 72 (1988), 221–226. |
| [3] | A. C. McIntosh, V. M. Gol'dshtein, I. Goldfarb, et al., Thermal explosion in a combustible gas containing fuel droplets, Combust. Th. Mod., 2 (1998), 153–165. |
| [4] | I. Goldfarb, V. M. Gol'dshtein, D. Katz, et al., Radiation effect on thermal explosion in a gas containing evaporating fuel droplets, Int. J. Ther. Sci., 46 (2007), 358–370. |
| [5] | M. R. Roussel and S. J. Fraser, Geometry of the steady-state approximation: perturbation and accelerated convergence methods, J. Chem. Phys., 93 (1990), 1072–1081. |
| [6] | M. R. Roussel and S. J. Fraser, Accurate steady-state approximations: implications for kinetics experiments and mechanism, J. Chem. Phys., 95 (1991), 8762–8770. |
| [7] | M. R. Roussel and S. J. Fraser, Global analysis of enzyme inhibition kinetics, textitJ. Chem. Phys., 97 (1993), 8316–8327. |
| [8] | M. R. Roussel and S. J. Fraser, Invariant manifold methods for metabolic model reduction, Chaos,196 (2001), 196–206. |
| [9] | A. Zagaris, H. G. Kaper and T. J. Kaper, Analysis of the computational singular perturbation reduction method for chemical kinetics, J. Non. Sci., 14 (2004), 59–91. |
| [10] | A. Zagaris, H. G. Kaper and T. J. Kaper, Fast and slow dynamics for the computational singular perturbation method, Soc. Indust. App. Math., 2 (2004), 613–638. |
| [11] | N. Berglunda and B. Gentzd, Geometric singular perturbation theory for stochastic differential equations, J. Diff. Eq., 191 (2003), 1–54. |
| [12] | N. Fenichel, Geometric singular perturbation theory for ordinary differential equations, J. Diff. Eq., 31 (1979), 53–98. |
| [13] | C. K. Jones, Geometric singular perturbation theory, 1609 of the series, Lec. Notes Math., Dyn. Syst. (2006), 44-118. |
| [14] | U. Maas and S. B. Pope, Implementation of simplified chemical kinetics based on intrinsic low-dimensional manifolds (PDF), Symposium (International) on Combustion, Twenty-Fourth Sym-posium on Combustion, (1992), 103–112. |
| [15] | U. Maas and S. B. Pope, Simplifying chemical kinetics: Intrinsic low-dimensional manifolds in composition space, Combust. Flame, 88 (1992), 239–264. |
| [16] | H. Bongers, J. A. Van Oijen and L. P. H. De Goey, Intrinsic low-dimensional manifold method extended with diffusion, Proc. Combust. Inst., 291 (2002), 1371–1378. |
| [17] | S. T. Alison, L. Whitehouse and L. Richard, The Estimation of Intrinsic Low Dimensional Man-ifold Dimension in Atmospheric Chemical Reaction Systems, Air Poll. Modell Simul., (2002), 245–263. |
| [18] | G. K. Hans and J. K. Tasso, Asymptotic analysis of two reduction methods for systems of chemical reactions, Phys. D, 165 (2002), 66–93. |
| [19] | V. Bykov, I. Goldfarb and V. Gol'dshtein, Singularly perturbed vector fields, J. Phys. Conf. Ser., 55 (2006), 28–44. |
| [20] | O. Nave, Singularly perturbed vector field method (SPVF) applied to combustion of monodisperse fuel spray, Diff. Eqs. Dyn. Syst., 27 (2018), 1–18. |
| [21] | M. Al-Tameemi, M. Chaplain and A. d'Onofrio, Evasion of tumours from the control of the im-mune system: consequences of brief encounters, Biol. Direct, 7 (2012), 1–22. |
| [22] | I. Kareva, F. Berezovskaya and C. Castillo-Chavez, Myeloid cells in tumour-immune interactions, J. Biol. Dyn., 4 (2010), 315–327. |
| [23] | L. Derbel, Analysis of a new model for tumor-immune system competition including long time scale effects, Math. Model Methods Appl. Sci., 14 (2004), 1657–1681. |
| [24] | K. E. Starkov and S. Bunimovich-Mendrazitsky, Dynamical properties and tumor clearance con-ditions for a nine-dimensional model of bladder cancer immunotherapy, Math. Biosci. Eng., 13 (2016), 1059–1075. |
| [25] | S. Bunimovich-Mendrazitsky, S. Halachmi and N. Kronik, Improving Bacillus Calmette-Guerin (BCG) immunotherapy for bladder cancer by adding interleukin 2 (IL−2): A mathematical model, Math. Med. Biol., 33 (2015), 1–30. |
| [26] | S. Brandau and H. Suttmann, Thirty years of BCG immunotherapy for non-muscle invasive blad-der cancer: a success story with room for improvement, Biomed. Pharmacother., 61 (2007), 299–305. |
| [27] | A. N. Tikhonov, Systems of differential equations containing small parameters multiplying the derivatives. Mat. Sborn., 31 (1952), 575–586. |
| [28] | R. Mahendran, Bacillus Calmette-Guerin immunotherapy-increasing dose as a means of improv-ing therapy?, Trans. Cancer Res., 6 (2017), 168–173. |
| [29] | A. Kiselyov, S. Bunimovich-Mendrazitsky and V. Startsev, Treatment of non-muscle invasive blad-der cancer with Bacillus Calmette-Guerin (BCG): biological markers and simulation studies, BBA Clin., 10 (2015), 27–34. |
| [30] | C. Pettenati and M. A.Ingersoll, Mechanisms of BCG immunotherapy and its outlook for bladder cancer, Nat. Rev. Urol., 15 (2018), 615–625. |
| [31] | A. H. Kitamur and T. Tsukamoto, Immunotherapy for urothelial carcinoma, Current status and perspectives, Cancers, 29 (2011), 3055–3072. |
| [32] | C. Yee, J. A. Thompson, D. Byrd, et al., Adoptive T cell therapy using antigen-specific CD8+ T cell clones for the treatment of patients with metastatic melanoma: in vivo persistence, migration, and antitumor effect of transferred T cells, Proc. Natl. Acad. Sci. U S A, 10 (2002), 16168–16173. |
| [33] | N. Kronik, Y. Kogan, P. G. Schlegel, et al., Improving T-cell immunotherapy for melanoma through a mathematically motivated strategy: efficacy in numbers?, J. Immunother., 35 (2012), 116–124. |
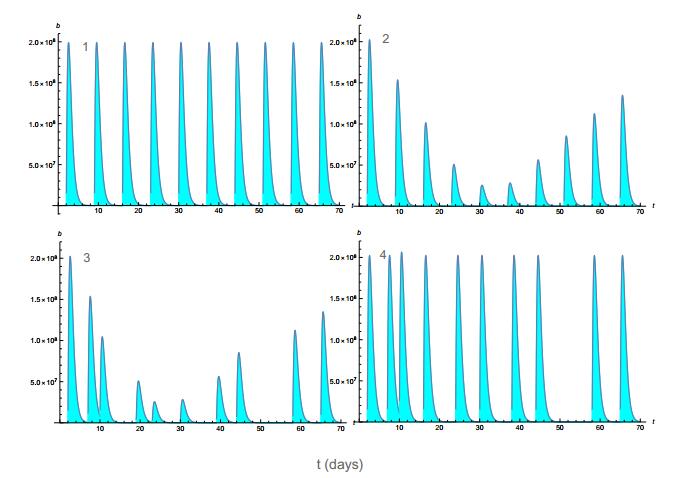
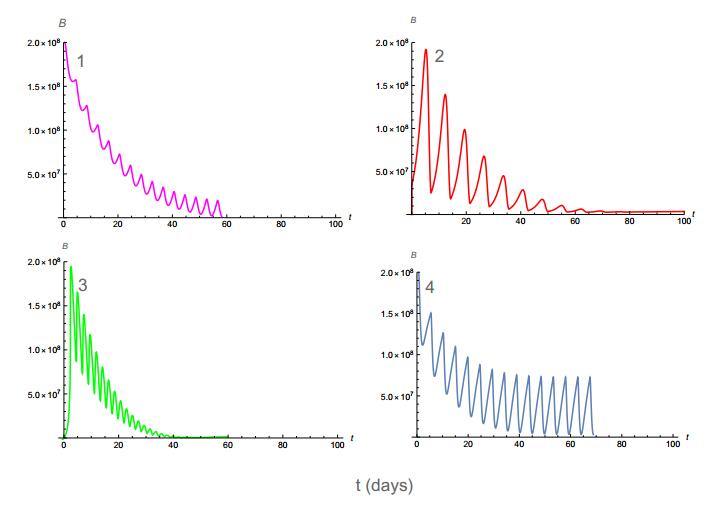
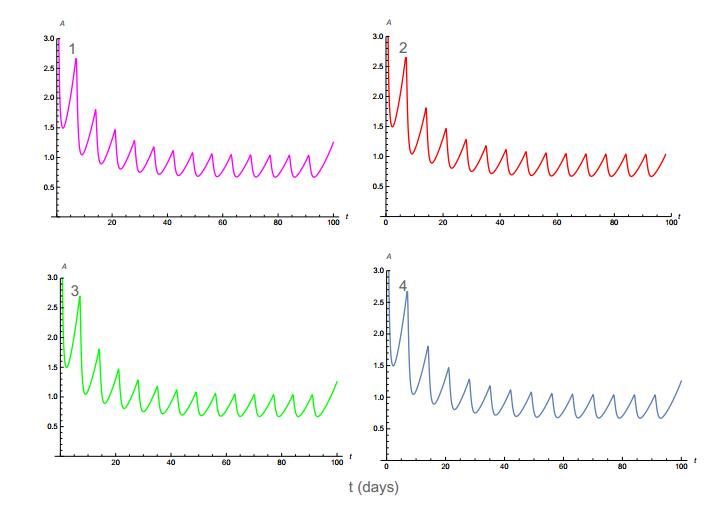
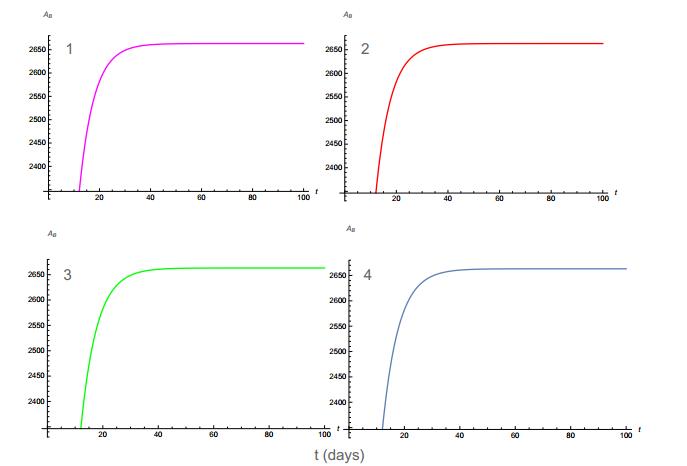
Figures(11) / Tables(4)

OPhir Nave, Shlomo Hareli, Miriam Elbaz, Itzhak Hayim Iluz, Svetlana Bunimovich-Mendrazitsky. BCG and IL − 2 model for bladder cancer treatment with fast and slow dynamics based on SPVF method—stability analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5346-5379. doi: 10.3934/mbe.2019267







 DownLoad:
DownLoad: