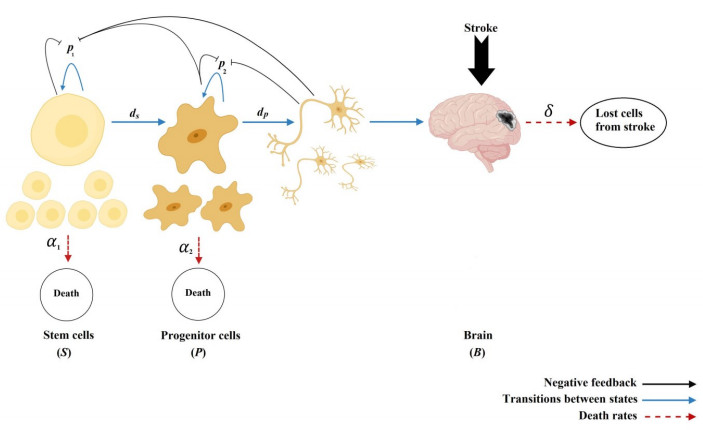
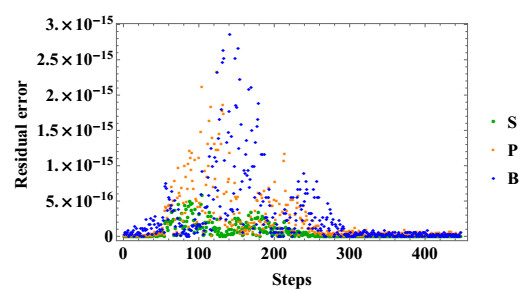
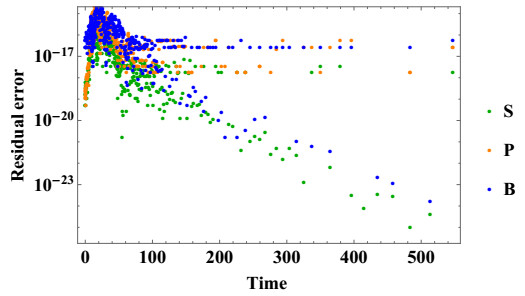
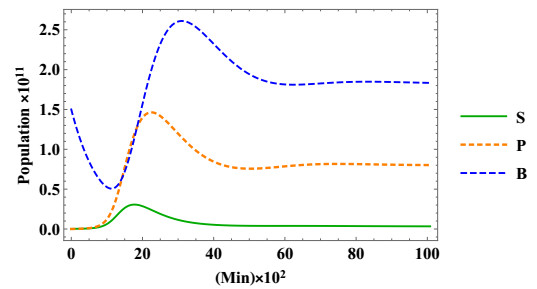
Stem cells play a critical role in regulatory operations, overseeing tissue regeneration and tissue homeostasis. In this paper, a mathematical model is proposed and analyzed to study the impact of stem cell transplantation on the dynamical behavior of stroke therapy, which is assumed to be based on transplanting dead brain cells following a stroke. We transform the method of using hierarchical cell systems into a method of using different compartment variables by using ordinary differential equations, each of which elucidates a well-defined differentiation stage along with the effect of mature cells in improving the brain function after a stroke. Stem cells, progenitor cells, and the impacts of the stem cells transplanted on brain cells are among the variables considered. The model is studied analytically and solved numerically using the fourth-order Runge-Kutta method. We analyze the structure of equilibria, the ability of neural stem cells to proliferate and differentiate, and the stability properties of equilibria for stem cell transplantation. The model is considered to be stable after transplantation if the stem cells and progenitor cells differentiate into mature nerve cells in the brain. The results of the model analysis and simulation facilitate the identification of various biologically probable parameter sets that can explain the optimal time for stem cell replacement of damaged brain cells. Associating the classified parameter sets with recent experimental and clinical findings contributes to a better understanding of therapeutic mechanisms that promote the reconstitution of brain cells after an ischemic stroke.
Citation: Awatif Jahman Alqarni, Azmin Sham Rambely, Sana Abdulkream Alharbi, Ishak Hashim. Dynamic behavior and stabilization of brain cell reconstitution after stroke under the proliferation and differentiation processes for stem cells[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6288-6304. doi: 10.3934/mbe.2021314
Stem cells play a critical role in regulatory operations, overseeing tissue regeneration and tissue homeostasis. In this paper, a mathematical model is proposed and analyzed to study the impact of stem cell transplantation on the dynamical behavior of stroke therapy, which is assumed to be based on transplanting dead brain cells following a stroke. We transform the method of using hierarchical cell systems into a method of using different compartment variables by using ordinary differential equations, each of which elucidates a well-defined differentiation stage along with the effect of mature cells in improving the brain function after a stroke. Stem cells, progenitor cells, and the impacts of the stem cells transplanted on brain cells are among the variables considered. The model is studied analytically and solved numerically using the fourth-order Runge-Kutta method. We analyze the structure of equilibria, the ability of neural stem cells to proliferate and differentiate, and the stability properties of equilibria for stem cell transplantation. The model is considered to be stable after transplantation if the stem cells and progenitor cells differentiate into mature nerve cells in the brain. The results of the model analysis and simulation facilitate the identification of various biologically probable parameter sets that can explain the optimal time for stem cell replacement of damaged brain cells. Associating the classified parameter sets with recent experimental and clinical findings contributes to a better understanding of therapeutic mechanisms that promote the reconstitution of brain cells after an ischemic stroke.

| [1] | A. Bergmann, H. Steller, Apoptosis, stem cells, and tissue regeneration, Sci. Signal, 145 (2010), 8. |
| [2] | Y. Nakata, P. Getto, A. Marciniak-Czochra, T. AlarcÃn, Stability analysis of multi-compartment models for cell production systems, Biol. Dyn., 6 (2012), 2–18. |
| [3] | F. Sa, L. Cf, C. Sk, Stem cell transplantation in Malaysia and future directions, Med. J. Malaysia, 63 (2008), 279–280. |
| [4] |
L. Wei, Z. Z. Wei, M. Q. Jiang, O. Mohamad, S. P. Yu, Stem cell transplantation therapy for multifaceted therapeutic benefits after stroke, Prog. Neurobiol., 157 (2017), 49–78. doi: 10.1016/j.pneurobio.2017.03.003

|
| [5] |
F. M. Watt, R. R. Driskell, The therapeutic potential of stem cells, Philos. Trans. Roy. Soc. London Ser. B, 365 (2010), 155–163. doi: 10.1098/rstb.2009.0149
|
| [6] | L. Hao, Z. Zou, H. Tian, Y. Zhang, H. Zhou, L. Liu, Stem cell-based therapies for ischemic stroke, Bio. Med. Res. Int., (2014), 468748. |
| [7] |
A. Benedek, D. Cernica, A. Mester, D. Opincariu, R. Hodas, I. Rodean, et al., Modern concepts in regenerative therapy for ischemic stroke: from stem cells for promoting angiogenesis to 3D-bioprinted scaffolds customized via carotid shear stress analysis, Int. J. Mol. Sci., 20 (2019), 2574. doi: 10.3390/ijms20102574
|
| [8] |
S. Beccari, J. Valero, M. Maletic-Savatic, A. Sierra, A simulation model of neuroprogenitor proliferation dynamics predicts age-related loss of hippocampal neurogenesis but not astrogenesis, Sci. Rep., 7 (2017), 16528. doi: 10.1038/s41598-017-16466-3
|
| [9] |
K. L. Spalding, O. Bergmann, K. Alkass, S. Bernard, M. Salehpour, H. Huttner, Dynamics of hippocampal neurogenesis in adult humans, Cell, 153 (2013), 1219–1227. doi: 10.1016/j.cell.2013.05.002
|
| [10] | D. Purves, G. J. Augustine, D. Fitzpatrick, L. C. Katz, A. S. LaMantia, J. O. McNamara, et al., Generation of neurons in the adult brain, Neuroscience, 364 (2001), 846–848. |
| [11] |
A. Arvidsson, T. Collin, D. Kirik, Z. Kokaia, O. Lindvall, Neuronal replacement from endogenous precursors in the adult brain after stroke, Nat. Med., 8 (2002), 963–970. doi: 10.1038/nm747
|
| [12] |
O. Y. Bang, E. H. Kim, J. M. Cha, G. J. Moon, Adult stem cell therapy for stroke: challenges and progress, J. Stroke, 18 (2016), 256–266. doi: 10.5853/jos.2016.01263
|
| [13] |
M. Chau, T. C. Deveau, M. Song, X. Gu, D. Chen, L. Wei, iPS cell transplantation increases regeneration and functional recovery after ischemic stroke in neonatal rats, Stem Cells, 32 (2014), 3075–3087. doi: 10.1002/stem.1802
|
| [14] |
A. C. Boese, Q. S. E. Le, D. Pham, M. H. Hamblin, J. P. Lee, Neural stem cell therapy for subacute and chronic ischemic stroke, Stem Cell Res. Ther., 9 (2018), 154. doi: 10.1186/s13287-018-0913-2
|
| [15] | E. W. Baker, H. A. Kinder, F. D. West, Neural stem cell therapy for stroke: A multimechanistic approach to restoring neurological function, Brain Behav., 3 (2019), 3. |
| [16] | E. Bible, D. Y. S. Chau, M. R. Alexander, J. Price, K. M. Shakesheff, M. Modo, The support of neural stem cells transplanted into stroke-induced brain cavities by PLGA particles, Biomaterials, 16 (2009), 2985–2994. |
| [17] |
S. M. Ziaee, P. Tabeshmehr, K. H. Haider, M. Farrokhi, A. Shariat, A. Amiri, et al., Optimization of time for neural stem cells transplantation for brain stroke in rats, Stem Cell Invest., 4 (2017), 29. doi: 10.21037/sci.2017.03.10
|
| [18] | N. A. Mn, S. A. Sj, N. A. A. Mn, A. S. Saperi, Direct medical cost of stroke: findings from a tertiary hospital in Malaysia, Med. J. Malaysia, 67 (2012), 473–477. |
| [19] | H. Bb, R. Aa, Risk factors and complications of acute ischaemic stroke patients at Hospital Universiti Kebangsaan Malaysia (HUKM), Med. J. Malaysia., 58 (2003), 499–505. |
| [20] | B. Z. Barkho, X. Zhao, Adult neural stem cells: response to stroke injury and potential for therapeutic applications, Cell Res. Ther., 6 (2011), 327–338. |
| [21] |
J. D. Jordan, W. J. Powers, Cerebral autoregulation and acute ischemic stroke, Am. J. Hypertens., 25 (2012), 946–950. doi: 10.1038/ajh.2012.53
|
| [22] |
S. Haas, N. Weidner, J. Winkler, Adult stem cell therapy in stroke, Curr. Opin. Neural, 18 (2005), 59–64. doi: 10.1097/00019052-200502000-00012
|
| [23] |
F. Wang, H. Tang, J. Zhu, J. H. Zhang, Transplanting mesenchymal stem cells for treatment of ischemic stroke, Cell Transplant., 27 (2018), 1825–1834. doi: 10.1177/0963689718795424
|
| [24] | C. D. Russo, A mathematical model of inflammation during ischemic stroke, Math. Model. Med., 30 (2010), 15–33. |
| [25] |
M. A. Dronne, J. P. Boissel, E. Grenier, H. Gilquin, M. Cucherat, M. Hommel, Mathematical modelling of an ischemic stroke: an integrative approach, Acta Biotheor., 52 (2004), 255–272. doi: 10.1023/B:ACBI.0000046597.53669.ff
|
| [26] | T. Lelekov-Boissard, G. C. Boissel, E. Grenier, M. A. Dronne, Exploration of beneficial and deleterious effects of inflammation in stroke: dynamics of inflammation cells, Philos. Trans. Roy. Soc. A., 367 (2009), 4699–4716. |
| [27] | M. E. Hall, N. K. Mohtaram, S. M. Willerth, R. Edwards, Modeling the behavior of human induced pluripotent stem cells seeded on melt electrospun scaffolds, J. Biomed. Eng., 11 (2017), 11–38. |
| [28] |
T. Stiehl, A. K. Marciniak-Czochra, Characterization of stem cells using mathematical models of multistage cell lineages, Math. Comput. Modell., 53 (2011), 1505–1517. doi: 10.1016/j.mcm.2010.03.057
|
| [29] |
T. Stiehl, A. Marciniak-Czochra, How to characterize stem cells? Contributions from mathematical modeling, Stem Cell Rep., 5 (2019), 57–65. doi: 10.1007/s40778-019-00155-0
|
| [30] |
F. Ziebell, A. Martin-Villalba, A. Marciniak-Czochra, Mathematical modelling of adult hippocampal neurogenesis: effects of altered stem cell dynamics on cell counts and bromodeoxyuridine-labelled cells, J. R. Soc. Interf., 11 (2014), 20140144. doi: 10.1098/rsif.2014.0144
|
| [31] |
J. M. Ashbourn, J. J. Miller, V. Reumers, V. Baekelandt, L. Geris, A mathematical model of adult subventricular neurogenesis, J. R. Soc. Interf., 9 (2012), 2414–2423. doi: 10.1098/rsif.2012.0193
|
| [32] |
A. J. Alqarni, A. S. Rambely, I. Hashim, Dynamic modelling of interactions between microglia and endogenous neural stem cells in the brain during a stroke, Mathematics, 8 (2020), 132. doi: 10.3390/math8101793
|
| [33] |
A. J. Alqarni, A. S. Rambely, I. Hashim, Dynamical simulation of effective stem cell transplantation for modulation of microglia responses in stroke treatment, Symmetry, 13 (2021), 404. doi: 10.3390/sym13040537
|
| [34] |
W. C. Lo, C. S. Chou, K. K. Gokoffski, F. Y. Wan, A. D. Lander, A. L. Calof, Q. Nie, Feedback regulation in multistage cell lineages, Math. Biosci. Eng., 6 (2009), 59–82. doi: 10.3934/mbe.2009.6.59
|
| [35] |
N. Omar, Y. Lokanathan, Z. R. M. Razi, R. B. H. Idrus, The effects of Centella asiatica (L.) Urban on neural differentiation of human mesenchymal stem cells in vitro, BMC Complement Altern. Med., 19 (2019), 167. doi: 10.1186/s12906-019-2581-x
|
| [36] |
M. X. Doss, A. Sachinidis, Current challenges of iPSC-based disease modeling and therapeutic implications, Cells, 8 (2019), 403. doi: 10.3390/cells8050403
|
| [37] |
R. Jin, G. Yang, G. Li, Inflammatory mechanisms in ischemic stroke: role of inflammatory cells, J. Leukoc. Biol., 87 (2010), 779–789. doi: 10.1189/jlb.1109766
|
| [38] |
S. E. Lakhan, A. Kirchgessner, M. Hofer, Inflammatory mechanisms in ischemic stroke: therapeutic approaches, J. Transl. Med., 7 (2009), 97. doi: 10.1186/1479-5876-7-97
|
| [39] |
T. Stiehl, A. Marciniak-Czochra, Stem cell self-renewal in regeneration and cancer: insights from mathematical modeling, Curr. Opin. Syst. Biol., 5 (2017), 112–120. doi: 10.1016/j.coisb.2017.09.006
|
| [40] | T. Stiehl, A. Marciniak-Czochra, Mathematical modelling of leukemogenesis and cancer stem cell dynamics, Math. Mod. Nat. Phenomena, 7 (2012), 7166–7202. |
| [41] |
M. S. Vieira, A. K. Santos, R. Vasconcellos, V. A. M. Goulart, R. C. Parreira, A. H. Kihara, Neural stem cell differentiation into mature neurons: Mechanisms of regulation and biotechnological applications, Biotechnol. Adv., 36 (2018), 1946–1970. doi: 10.1016/j.biotechadv.2018.08.002
|
| [42] |
M. K. Tobin, J. A. Bonds, R. D. Minshall, D. A. Pelligrino, F. D. Testai, O. Lazarov, Neurogenesis and inflammation after ischemic stroke: what is known and where we go from here, J. Cereb. Blood Flow Metab., 34 (2014), 1573–1584. doi: 10.1038/jcbfm.2014.130
|
| [43] |
F. A. Azevedo, L. R. Carvalho, L. T. Grinberg, J. M. Farfel, R. E. Ferretti, R. E. Leite et al., Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain, J. Comp. Neural, 513 (2009), 532–541. doi: 10.1002/cne.21974
|
| [44] |
S. Ryu, S. H. Lee, S. U. Kim, B. W. Yoon, Human neural stem cells promote proliferation of endogenous neural stem cells and enhance angiogenesis in ischemic rat brain, Neural Regen. Res., 11 (2016), 298–304. doi: 10.4103/1673-5374.177739
|
| [45] |
K. Oki, J. Tatarishvili, J. Wood, P. Koch, S. Wattananit, Y. Mine, et al., Human-induced pluripotent stem cells form functional neurons and improve recovery after grafting in stroke-damaged brain, Stem Cells, 30 (2012), 1120–1133. doi: 10.1002/stem.1104
|
Figures(4) / Tables(1)

Awatif Jahman Alqarni, Azmin Sham Rambely, Sana Abdulkream Alharbi, Ishak Hashim. Dynamic behavior and stabilization of brain cell reconstitution after stroke under the proliferation and differentiation processes for stem cells[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6288-6304. doi: 10.3934/mbe.2021314







 DownLoad:
DownLoad: