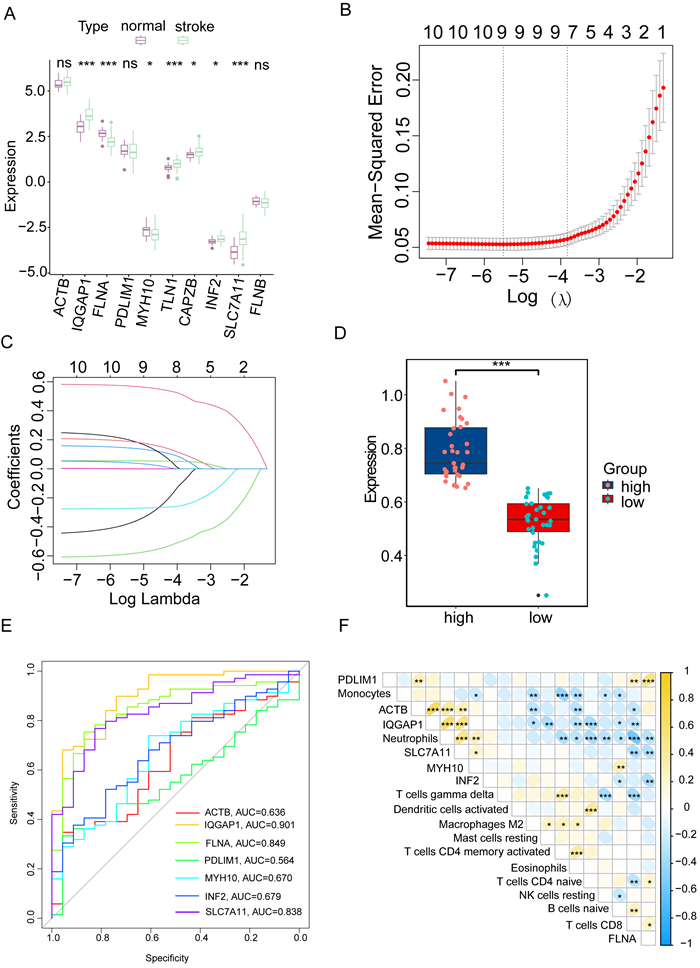
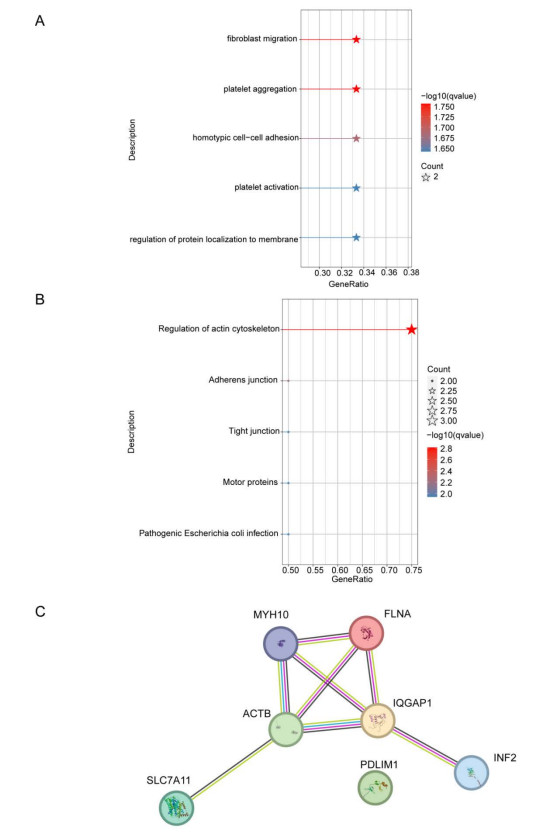
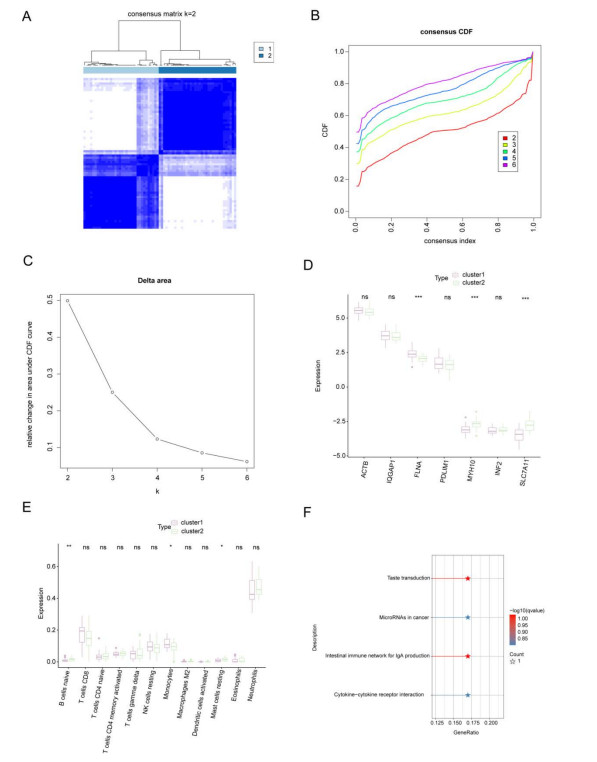
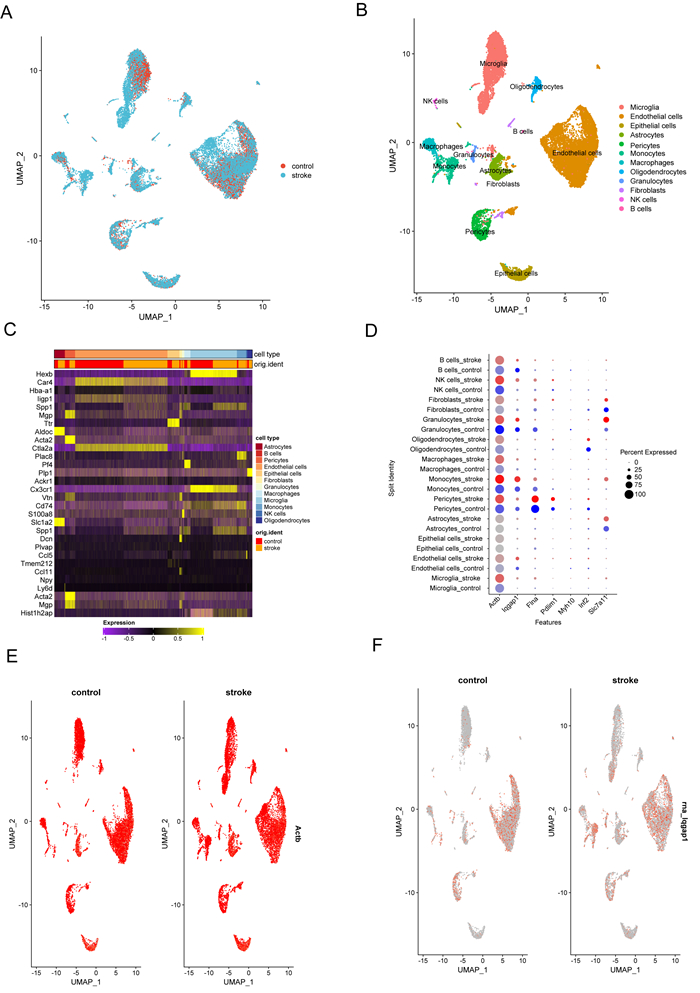
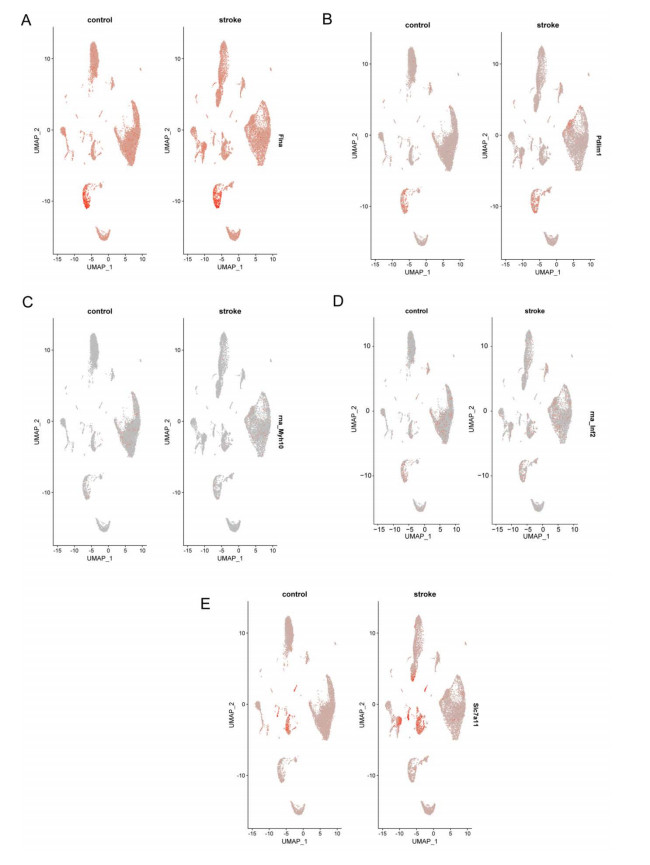
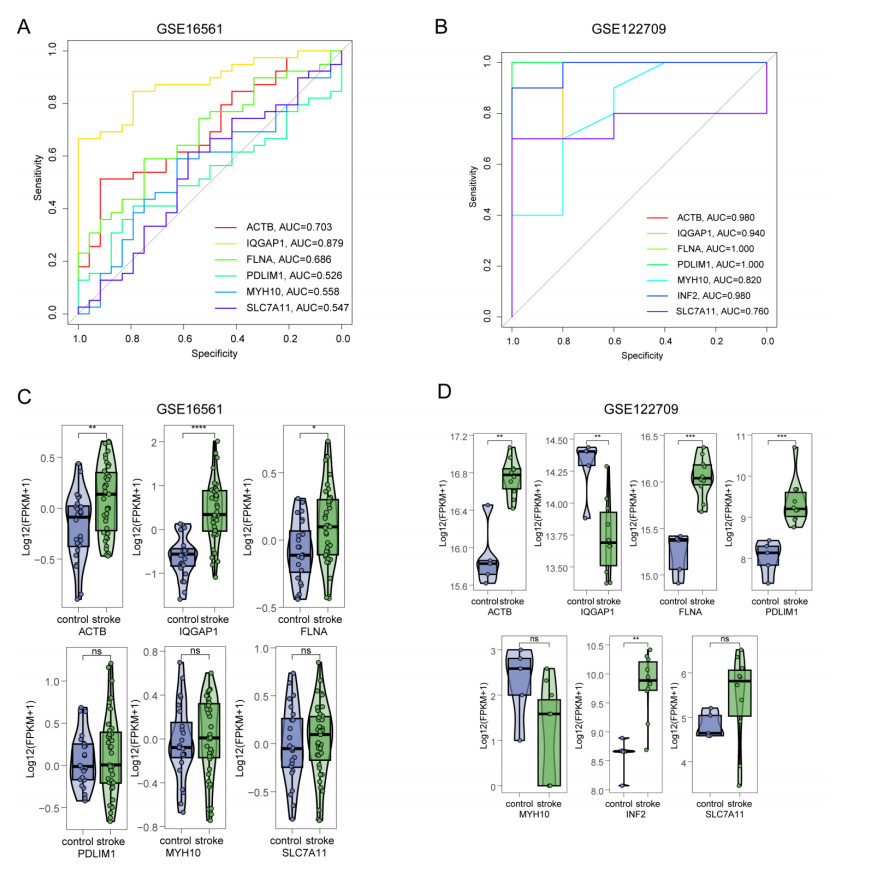
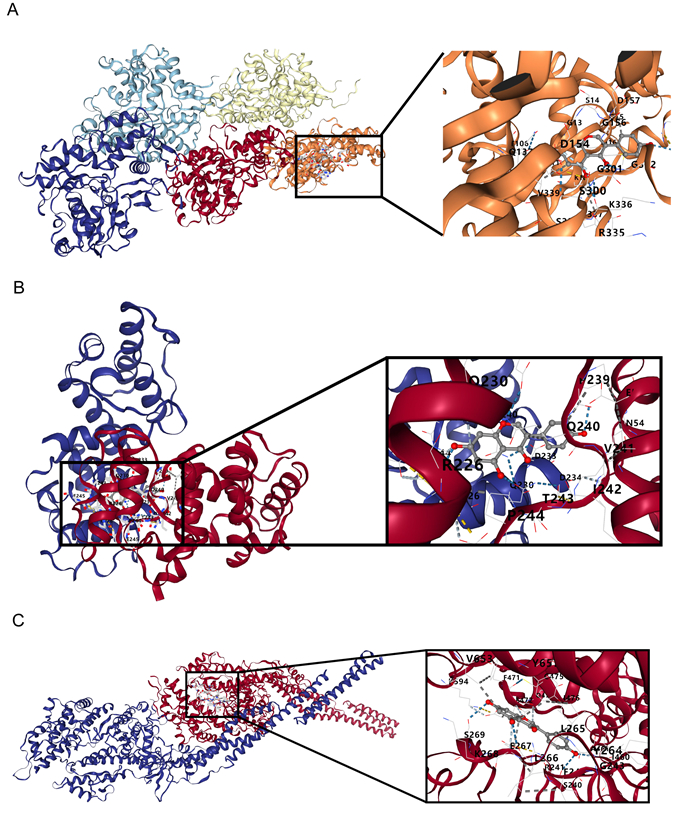
Immune infiltration plays a pivotal role in the pathogenesis of ischemic stroke. A novel form of cell death known as disulfidptosis has emerged in recent studies. However, there is currently a lack of research investigating the regulatory mechanism of disulfidptosis-related genes in immune infiltration during ischemic stroke. Using machine learning methods, we identified candidate key disulfidptosis-related genes (DRGs). Subsequently, we performed an analysis of immune cell infiltration to investigate the dysregulation of immune cells in the context of ischemic stroke. We assessed their diagnostic value by employing receiver operating characteristic (ROC) curves. To gain further insights, we conducted functional enrichment analyses to elucidate the signaling pathways associated with these seven DRGs. We identified two distinct subclusters based on the expression patterns of these seven DRGs. The unique roles of these subclusters were further evaluated through KEGG analysis and immune infiltration studies. Furthermore, we validated the expression profiles of these seven DRGs using both single-cell datasets and external datasets. Lastly, molecular docking was performed to explore potential drugs for the treatment of ischemic stroke. We identified seven DRGs. The seven DRGs are related to immune cells. Additionally, these seven DRGs also demonstrate potential diagnostic value in ischemic stroke. Functional enrichment analysis highlighted pathways such as platelet aggregation and platelet activation. Two subclusters related to disulfidptosis were defined, and functional enrichment analysis of their differentially expressed genes (DEGs) primarily involved pathways like cytokine-cytokine receptor interaction. Single-cell analysis indicated that these seven DRGs were primarily distributed among immune cell types. Molecular docking results suggested that genistein might be a potential therapeutic drug. This study has opened up new avenues for exploring the causes of ischemic stroke and developing potential therapeutic targets.
Citation: Rongxing Qin, Lijuan Huang, Wei Xu, Qingchun Qin, Xiaojun Liang, Xinyu Lai, Xiaoying Huang, Minshan Xie, Li Chen. Identification of disulfidptosis-related genes and analysis of immune infiltration characteristics in ischemic strokes[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18939-18959. doi: 10.3934/mbe.2023838
Immune infiltration plays a pivotal role in the pathogenesis of ischemic stroke. A novel form of cell death known as disulfidptosis has emerged in recent studies. However, there is currently a lack of research investigating the regulatory mechanism of disulfidptosis-related genes in immune infiltration during ischemic stroke. Using machine learning methods, we identified candidate key disulfidptosis-related genes (DRGs). Subsequently, we performed an analysis of immune cell infiltration to investigate the dysregulation of immune cells in the context of ischemic stroke. We assessed their diagnostic value by employing receiver operating characteristic (ROC) curves. To gain further insights, we conducted functional enrichment analyses to elucidate the signaling pathways associated with these seven DRGs. We identified two distinct subclusters based on the expression patterns of these seven DRGs. The unique roles of these subclusters were further evaluated through KEGG analysis and immune infiltration studies. Furthermore, we validated the expression profiles of these seven DRGs using both single-cell datasets and external datasets. Lastly, molecular docking was performed to explore potential drugs for the treatment of ischemic stroke. We identified seven DRGs. The seven DRGs are related to immune cells. Additionally, these seven DRGs also demonstrate potential diagnostic value in ischemic stroke. Functional enrichment analysis highlighted pathways such as platelet aggregation and platelet activation. Two subclusters related to disulfidptosis were defined, and functional enrichment analysis of their differentially expressed genes (DEGs) primarily involved pathways like cytokine-cytokine receptor interaction. Single-cell analysis indicated that these seven DRGs were primarily distributed among immune cell types. Molecular docking results suggested that genistein might be a potential therapeutic drug. This study has opened up new avenues for exploring the causes of ischemic stroke and developing potential therapeutic targets.

| [1] |
B. C. V. Campbell, P. Khatri, Stroke, Lancet (London, England), 396 (2020), 129–142. http://doi.org/10.1016/s0140-6736(20)31179-x doi: 10.1016/s0140-6736(20)31179-x

|
| [2] |
M. Katan, A. Luft, Global Burden of Stroke, Seminars Neurol., 38 (2018), 208–211. http://doi.org/10.1055/s-0038-1649503 doi: 10.1055/s-0038-1649503
|
| [3] |
V. L. Feigin, G. Nguyen, K. Cercy, C. O. Johnson, T. Alam, P. G. Parmar, et al., Global, regional, and country-specific lifetime risks of stroke, 1990 and 2016, New England J. Med., 379 (2018), 2429–2437. http://doi.org/10.1056/NEJMoa1804492 doi: 10.1056/NEJMoa1804492
|
| [4] |
S. S. Virani, A. Alonso, E. J. Benjamin, M. S. Bittencourt, C. W. Callaway, A. P. Carson, et al., Heart disease and stroke statistics-2020 update: A report from the American Heart Association, Circulation, 141 (2020), e139–e596. http://doi.org/10.1161/cir.0000000000000757 doi: 10.1161/cir.0000000000000757
|
| [5] |
A. K. Boehme, C. Esenwa, M. S. Elkind, Stroke Risk Factors, Genetics, and Prevention, Circulation research, 120 (2017), 472–495. http://doi.org/10.1161/circresaha.116.308398 doi: 10.1161/circresaha.116.308398
|
| [6] |
C. Gumbinger, B. Reuter, C. Stock, T. Sauer, H. Wiethölter, I. Bruder, et al., Time to treatment with recombinant tissue plasminogen activator and outcome of stroke in clinical practice: Retrospective analysis of hospital quality assurance data with comparison with results from randomised clinical trials, BMJ (Clinical Research ed), 348 (2014), g3429. http://doi.org/10.1136/bmj.g3429 doi: 10.1136/bmj.g3429
|
| [7] |
X. Liu, L. Nie, Y. Zhang, Y. Yan, C. Wang, M. Colic, et al., Actin cytoskeleton vulnerability to disulfide stress mediates disulfidptosis, Nat. Cell Biol., 25 (2023), 404–414. http://doi.org/10.1038/s41556-023-01091-2 doi: 10.1038/s41556-023-01091-2
|
| [8] |
N. Maehara, K. Taniguchi, A. Okuno, H. Ando, A. Hirota, Z. Li, et al., AIM/CD5L attenuates DAMPs in the injured brain and thereby ameliorates ischemic stroke, Cell Rep., 36 (2021), 109693. http://doi.org/10.1016/j.celrep.2021.109693 doi: 10.1016/j.celrep.2021.109693
|
| [9] |
H. Bektas, G. Vural, S. Gumusyayla, O. Deniz, M. Alisik, O. Erel, Dynamic thiol-disulfide homeostasis in acute ischemic stroke patients, Acta Neurol. Belg., 116 (2016), 489–494. http://doi.org/10.1007/s13760-016-0598-1 doi: 10.1007/s13760-016-0598-1
|
| [10] |
A. B. Yilmaz, S. Gokhan, A. Sener, O. Erel, Analysis of Neutrophil/Lymphocyte ratio and Thiol/Disulfide homeostasis parameters in patients admitted to the emergency department with ischemic stroke, Pakistan J Med. Sci., 34 (2018), 1418–1423. http://doi.org/10.12669/pjms.346.16242 doi: 10.12669/pjms.346.16242
|
| [11] | L. Wang, X. Zhang, X. Xiong, H. Zhu, R. Chen, S. Zhang, et al., Nrf2 regulates oxidative stress and its role in cerebral ischemic stroke, antioxidants (Basel, Switzerland), 11 (2022). http://doi.org/10.3390/antiox11122377 |
| [12] |
H. X. Chu, H. A. Kim, S. Lee, J. P. Moore, C. T. Chan, A. Vinh, et al., Immune cell infiltration in malignant middle cerebral artery infarction: Comparison with transient cerebral ischemia, J. Cerebral Blood Flow Metabol. Offic. J. Int. Soc. Cerebral Blood Flow Metabol., 34 (2014), 450–459. http://doi.org/10.1038/jcbfm.2013.217 doi: 10.1038/jcbfm.2013.217
|
| [13] |
Y. Ma, A. Yabluchanskiy, R. P. Iyer, P. L. Cannon, E. R. Flynn, M. Jung, et al., Temporal neutrophil polarization following myocardial infarction, Cardiovascular Res., 110 (2016), 51–61. http://doi.org/10.1093/cvr/cvw024 doi: 10.1093/cvr/cvw024
|
| [14] |
Z. Jian, R. Liu, X. Zhu, D. Smerin, Y. Zhong, L. Gu, et al., The Involvement and Therapy Target of Immune Cells After Ischemic Stroke, Frontiers Immunol., 10 (2019), 2167. http://doi.org/10.3389/fimmu.2019.02167 doi: 10.3389/fimmu.2019.02167
|
| [15] |
C. Iadecola, M. S. Buckwalter, J. Anrather, Immune responses to stroke: Mechanisms, modulation, and therapeutic potential, J. Clin. Invest., 130 (2020), 2777–2788. http://doi.org/10.1172/jci135530 doi: 10.1172/jci135530
|
| [16] |
B. Stamova, G. C. Jickling, B. P. Ander, X. Zhan, D. Liu, R. Turner, et al., Gene expression in peripheral immune cells following cardioembolic stroke is sexually dimorphic, PloS One, 9 (2014), e102550. http://doi.org/10.1371/journal.pone.0102550 doi: 10.1371/journal.pone.0102550
|
| [17] |
T. L. Barr, Y. Conley, J. Ding, A. Dillman, S. Warach, A. Singleton, et al., Genomic biomarkers and cellular pathways of ischemic stroke by RNA gene expression profiling, Neurology, 75 (2010), 1009–1014. http://doi.org/10.1212/WNL.0b013e3181f2b37f doi: 10.1212/WNL.0b013e3181f2b37f
|
| [18] |
W. Zhu, L. Tian, X. Yue, J. Liu, Y. Fu, Y. Yan, LncRNA expression profiling of ischemic stroke during the transition from the acute to subacute stage, Front. Neurol., 10 (2019), 36. http://doi.org/10.3389/fneur.2019.00036 doi: 10.3389/fneur.2019.00036
|
| [19] |
K. Zheng, L. Lin, W. Jiang, L. Chen, X. Zhang, Q. Zhang, et al., Single-cell RNA-seq reveals the transcriptional landscape in ischemic stroke, J. Cerebral Blood Flow Metabol. Offic. J. Int. Soc. Cerebral Blood Flow Metabol., 42 (2022), 56–73. http://doi.org/10.1177/0271678x211026770 doi: 10.1177/0271678x211026770
|
| [20] |
S. Engebretsen, J. Bohlin, Statistical predictions with glmnet, Clin. Epigenet., 11 (2019), 123. http://doi.org/10.1186/s13148-019-0730-1 doi: 10.1186/s13148-019-0730-1
|
| [21] |
X. Robin, N. Turck, A. Hainard, N. Tiberti, F. Lisacek, J. C. Sanchez, et al., pROC: An open-source package for R and S+ to analyze and compare ROC curves, BMC Bioinform., 12 (2011), 77. http://doi.org/10.1186/1471-2105-12-77 doi: 10.1186/1471-2105-12-77
|
| [22] |
M. D. Wilkerson, D. N. Hayes, ConsensusClusterPlus: A class discovery tool with confidence assessments and item tracking, Bioinformatics (Oxford, England), 26 (2010), 1572–1573. http://doi.org/10.1093/bioinformatics/btq170 doi: 10.1093/bioinformatics/btq170
|
| [23] |
A. Gribov, M. Sill, S. Lück, F. Rücker, K. Döhner, L. Bullinger, et al., SEURAT: Visual analytics for the integrated analysis of microarray data, BMC Med. Genom., 3 (2010), 21. http://doi.org/10.1186/1755-8794-3-21 doi: 10.1186/1755-8794-3-21
|
| [24] |
M. Yoo, J. Shin, J. Kim, K. A. Ryall, K. Lee, S. Lee, et al., DSigDB: Drug signatures database for gene set analysis, Bioinformatics (Oxford, England), 31 (2015), 3069–3071. http://doi.org/10.1093/bioinformatics/btv313 doi: 10.1093/bioinformatics/btv313
|
| [25] |
H. M. Berman, J. Westbrook, Z. Feng, G. Gilliland, T. N. Bhat, H. Weissig, et al., The Protein Data Bank, Nucleic Acids Res., 28 (2000), 235–242. http://doi.org/10.1093/nar/28.1.235 doi: 10.1093/nar/28.1.235
|
| [26] |
S. Kim, J. Chen, T. Cheng, A. Gindulyte, J. He, S. He, et al., PubChem in 2021: New data content and improved web interfaces, Nucleic Acids Res., 49 (2021), D1388–d1395. http://doi.org/10.1093/nar/gkaa971 doi: 10.1093/nar/gkaa971
|
| [27] |
Y. Liu, X. Yang, J. Gan, S. Chen, Z. X. Xiao, Y. Cao, CB-Dock2: Improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting, Nucleic Acids Res., 50 (2022), W159–w164. http://doi.org/10.1093/nar/gkac394 doi: 10.1093/nar/gkac394
|
| [28] |
S. F. Nabavi, M. Daglia, R. Tundis, M. R. Loizzo, E. Sobarzo-Sanchez, I. E. Orhan, et al., Genistein: A boon for mitigating ischemic stroke, Current Topics Med. Chem., 15 (2015), 1714–1721. http://doi.org/10.2174/1568026615666150427122709 doi: 10.2174/1568026615666150427122709
|
| [29] |
S. K. Feske, Ischemic Stroke, Am. J. Med., 134 (2021), 1457–1464. http://doi.org/10.1016/j.amjmed.2021.07.027 doi: 10.1016/j.amjmed.2021.07.027
|
| [30] |
R. Kudoh, K. Mikami, M. Kitajima, K. Aizu, Y. Kitajima, J. Hagii, et al., Spontaneous micro-aggregation of platelets predicts clinical outcome in acute ischemic stroke, J. Stroke Cerebrovascular Diseases Official J. Nat. Stroke Assoc., 27 (2018), 2074–2081. http://doi.org/10.1016/j.jstrokecerebrovasdis.2018.03.004 doi: 10.1016/j.jstrokecerebrovasdis.2018.03.004
|
| [31] |
A. M. Kollikowski, M. Pham, A. G. März, L. Papp, B. Nieswandt, G. Stoll, et al., Platelet activation and chemokine release are related to local neutrophil-dominant inflammation during hyperacute human stroke, Translat. Stroke Res., 13 (2022), 364–369. http://doi.org/10.1007/s12975-021-00938-w doi: 10.1007/s12975-021-00938-w
|
| [32] |
L. Li, L. Dong, Z. Xiao, W. He, J. Zhao, H. Pan, et al., Integrated analysis of the proteome and transcriptome in a MCAO mouse model revealed the molecular landscape during stroke progression, J. Adv. Res., 24 (2020), 13–27. http://doi.org/10.1016/j.jare.2020.01.005 doi: 10.1016/j.jare.2020.01.005
|
| [33] |
L. Artman, V. Dormoy-Raclet, C. von Roretz, I. E. Gallouzi, Planning your every move: The role of β-actin and its post-transcriptional regulation in cell motility, Seminars Cell Develop. Biol., 34 (2014), 33–43. http://doi.org/10.1016/j.semcdb.2014.05.012 doi: 10.1016/j.semcdb.2014.05.012
|
| [34] |
G. Chen, Y. Zou, X. Zhang, L. Xu, Q. Hu, T. Li, et al., β-Actin protein expression differs in the submandibular glands of male and female mice, Cell Biol. Int., 40 (2016), 779–786. http://doi.org/10.1002/cbin.10613 doi: 10.1002/cbin.10613
|
| [35] | I. M. Herman, Actin isoforms, Current opinion in cell biology, 5 (1993), 48–55. http://doi.org/10.1016/s0955-0674(05)80007-9 |
| [36] |
M. Moustafa-Bayoumi, M. A. Alhaj, O. El-Sayed, S. Wisel, M. A. Chotani, Z. A. Abouelnaga, et al., Vascular hypertrophy and hypertension caused by transgenic overexpression of profilin 1, J. Biol. Chem., 282 (2007), 37632–37639. http://doi.org/10.1074/jbc.M703227200 doi: 10.1074/jbc.M703227200
|
| [37] |
J. C. Nelson, A. K. Stavoe, D. A. Colón-Ramos, The actin cytoskeleton in presynaptic assembly, Cell Adhes. Migr., 7 (2013), 379–387. http://doi.org/10.4161/cam.24803 doi: 10.4161/cam.24803
|
| [38] |
S. Yang, Y. Zhao, X. Chen, X. Lu, Y. Chen, X. Zhao, et al., The ACTB Variants and Alcohol Drinking Confer Joint Effect to Ischemic Stroke in Chinese Han Population, J. Atheroscl. Thromb., 27 (2020), 226–244. http://doi.org/10.5551/jat.49536 doi: 10.5551/jat.49536
|
| [39] |
A. M. Abel, K. M. Schuldt, K. Rajasekaran, D. Hwang, M. J. Riese, S. Rao, et al., IQGAP1: Insights into the function of a molecular puppeteer, Molecular Immun., 65 (2015), 336–349. http://doi.org/10.1016/j.molimm.2015.02.012 doi: 10.1016/j.molimm.2015.02.012
|
| [40] |
Y. Liu, W. Liang, Q. Yang, Z. Ren, X. Chen, D. Zha, et al., IQGAP1 mediates angiotensin Ⅱ-induced apoptosis of podocytes via the ERK1/2 MAPK signaling pathway, Am. J. Nephrol., 38 (2013), 430–444. http://doi.org/10.1159/000355970 doi: 10.1159/000355970
|
| [41] |
A. Sato, T. Naito, A. Hiramoto, K. Goda, T. Omi, Y. Kitade, et al., Association of RNase L with a Ras GTPase-activating-like protein IQGAP1 in mediating the apoptosis of a human cancer cell-line, FEBS J., 277 (2010), 4464–4473. http://doi.org/10.1111/j.1742-4658.2010.07833.x doi: 10.1111/j.1742-4658.2010.07833.x
|
| [42] |
D. Xu, T. Kong, Z. Shao, M. Liu, R. Zhang, S. Zhang, et al., Orexin-A alleviates astrocytic apoptosis and inflammation via inhibiting OX1R-mediated NF-κB and MAPK signaling pathways in cerebral ischemia/reperfusion injury, Biochimica et biophysica acta Molecular basis of disease, 1867 (2021), 166230. http://doi.org/10.1016/j.bbadis.2021.166230 doi: 10.1016/j.bbadis.2021.166230
|
| [43] |
B. Xu, J. Xu, N. Cai, M. Li, L. Liu, Y. Qin, et al., Roflumilast prevents ischemic stroke-induced neuronal damage by restricting GSK3β-mediated oxidative stress and IRE1α/TRAF2/JNK pathway, Free Rad. Biol. Med., 163 (2021), 281–296. http://doi.org/10.1016/j.freeradbiomed.2020.12.018 doi: 10.1016/j.freeradbiomed.2020.12.018
|
| [44] |
S. Bandaru, C. Ala, R. Salimi, M. K. Akula, M. Ekstrand, S. Devarakonda, et al., Targeting filamin a reduces macrophage activity and atherosclerosis, Circulation, 140 (2019), 67–79. http://doi.org/10.1161/circulationaha.119.039697 doi: 10.1161/circulationaha.119.039697
|
| [45] |
T. Hasegawa, K. Ohno, S. Funahashi, K. Miyazaki, A. Nagano, K. Sato, CLP36 interacts with palladin in dorsal root ganglion neurons, Neurosci. Letters, 476 (2010), 53–57. http://doi.org/10.1016/j.neulet.2010.03.081 doi: 10.1016/j.neulet.2010.03.081
|
| [46] |
Y. W. Jun, S. Lee, B. K. Ban, J. A. Lee, F. B. Gao, Non-muscle MYH10/myosin ⅡB recruits ESCRT-Ⅲ to participate in autophagosome closure to maintain neuronal homeostasis, Autophagy, (2023), 1–17. http://doi.org/10.1080/15548627.2023.2169309 doi: 10.1080/15548627.2023.2169309
|
| [47] |
B. Calabrese, S. L. Jones, Y. Shiraishi-Yamaguchi, M. Lingelbach, U. Manor, T. M. Svitkina, et al., INF2-mediated actin filament reorganization confers intrinsic resilience to neuronal ischemic injury, Nat. Commun., 13 (2022), 6037. http://doi.org/10.1038/s41467-022-33268-y doi: 10.1038/s41467-022-33268-y
|
| [48] |
Y. Yuan, Y. Zhai, J. Chen, X. Xu, H. Wang, Kaempferol ameliorates oxygen-glucose deprivation/reoxygenation-induced neuronal ferroptosis by activating Nrf2/SLC7A11/GPX4 axis, Biomolecules, 11 (2021). http://doi.org/10.3390/biom11070923 doi: 10.3390/biom11070923
|
| [49] |
X. Guan, Z. Li, S. Zhu, M. Cheng, Y. Ju, L. Ren, et al., Galangin attenuated cerebral ischemia-reperfusion injury by inhibition of ferroptosis through activating the SLC7A11/GPX4 axis in gerbils, Life Sci., 264 (2021), 118660. http://doi.org/10.1016/j.lfs.2020.118660 doi: 10.1016/j.lfs.2020.118660
|
| [50] |
J. Anrather, C. Iadecola, Inflammation and Stroke: An Overview, Neurother. J. Am. Soc. Exper. Neurotherapeut., 13 (2016), 661–670. http://doi.org/10.1007/s13311-016-0483-x doi: 10.1007/s13311-016-0483-x
|
| [51] |
A. Rayasam, M. Hsu, J. A. Kijak, L. Kissel, G. Hernandez, M. Sandor, et al., Immune responses in stroke: how the immune system contributes to damage and healing after stroke and how this knowledge could be translated to better cures?, Immunology, 154 (2018), 363–376. http://doi.org/10.1111/imm.12918 doi: 10.1111/imm.12918
|
| [52] |
M. G. Manz, S. Boettcher, Emergency granulopoiesis, Nature Rev. Immunol., 14 (2014), 302–314. http://doi.org/10.1038/nri3660 doi: 10.1038/nri3660
|
| [53] |
J. Xue, W. Huang, X. Chen, Q. Li, Z. Cai, T. Yu, et al., Neutrophil-to-Lymphocyte ratio is a prognostic marker in acute ischemic stroke, J. Stroke Cerebrovascular Diseases Offic. J. Nat. Stroke Assoc., 26 (2017), 650–657. http://doi.org/10.1016/j.jstrokecerebrovasdis.2016.11.010 doi: 10.1016/j.jstrokecerebrovasdis.2016.11.010
|
| [54] |
H. X. Chu, B. R. Broughton, H. A. Kim, S. Lee, G. R. Drummond, C. G. Sobey, Evidence that Ly6C(hi) monocytes are protective in acute ischemic stroke by promoting M2 macrophage polarization, Stroke, 46 (2015), 1929–1937. http://doi.org/10.1161/strokeaha.115.009426 doi: 10.1161/strokeaha.115.009426
|
| [55] |
A. Smirkin, H. Matsumoto, H. Takahashi, A. Inoue, M. Tagawa, S. Ohue, et al., Iba1(+)/NG2(+) macrophage-like cells expressing a variety of neuroprotective factors ameliorate ischemic damage of the brain, J. Cerebral Blood Flow Metabol. Offic. J. Int. Soc. Cerebral Blood Flow. Metabol., 30 (2010), 603–615. http://doi.org/10.1038/jcbfm.2009.233 doi: 10.1038/jcbfm.2009.233
|
| [56] |
J. Pedragosa, F. Miró-Mur, A. Otxoa-de-Amezaga, C. Justicia, F. Ruíz-Jaén, P. Ponsaerts, et al., CCR2 deficiency in monocytes impairs angiogenesis and functional recovery after ischemic stroke in mice, J. Cerebral Blood Flow Metabol. Offic. J. Int. Soc. Cerebral Blood Flow. Metabol., 40 (2020), S98–s116. http://doi.org/10.1177/0271678x20909055 doi: 10.1177/0271678x20909055
|
| [57] | U. M. Selvaraj, A. M. Stowe, Long-term T cell responses in the brain after an ischemic stroke, Discovery Med., 24 (2017), 323–333. |
| [58] |
D. Klebe, D. McBride, J. J. Flores, J. H. Zhang, J. Tang, Modulating the immune response towards a neuroregenerative peri-injury milieu after cerebral hemorrhage, J. Neuroimmune Pharm. Offic. J. Soc. Neuroimmune Pharmacol., 10 (2015), 576–586. http://doi.org/10.1007/s11481-015-9613-1 doi: 10.1007/s11481-015-9613-1
|
| [59] |
L. Peruzzotti-Jametti, M. Donegá, E. Giusto, G. Mallucci, B. Marchetti, S. Pluchino, The role of the immune system in central nervous system plasticity after acute injury, Neuroscience, 283 (2014), 210–221. http://doi.org/10.1016/j.neuroscience.2014.04.036 doi: 10.1016/j.neuroscience.2014.04.036
|
| [60] | L. B. Guo, S. Liu, F. Zhang, G. S. Mao, L. Z. Sun, Y. Liu, The role of eosinophils in stroke: A pilot study, European Rev. Med. Pharm. Sci., 19 (2015), 3643–3648. |
| [61] |
Y. Gan, Q. Liu, W. Wu, J. X. Yin, X. F. Bai, R. Shen, et al., Ischemic neurons recruit natural killer cells that accelerate brain infarction, Proceed. Nat. Aca. Sci. USA, 111 (2014), 2704–2709. http://doi.org/10.1073/pnas.1315943111 doi: 10.1073/pnas.1315943111
|
| [62] |
S. Xu, J. Lu, A. Shao, J. H. Zhang, J. Zhang, Glial cells: Role of the immune response in ischemic stroke, Frontiers Immunol., 11 (2020), 294. http://doi.org/10.3389/fimmu.2020.00294 doi: 10.3389/fimmu.2020.00294
|
| [63] | Z. Banecka-Majkutewicz, W. Sawuła, L. Kadziński, A. Węgrzyn, B. Banecki, Homocysteine, heat shock proteins, genistein and vitamins in ischemic stroke--pathogenic and therapeutic implications, Acta Bioch. Polon., 59 (2012), 495–499. |
| [64] | M. Petras, Z. Tatarkova, M. Kovalska, D. Mokra, D. Dobrota, J. Lehotsky, et al., Hyperhomocysteinemia as a risk factor for the neuronal system disorders, J. Physiol. Pharm. Offic. J. Polish Physiol. Soc., 65 (2014), 15–23. |
| [65] |
Z. Banecka-Majkutewicz, L. Kadziński, M. Grabowski, S. Bloch, R. Kaźmierkiewicz, J. Jakóbkiewicz-Banecka, et al., Evidence for interactions between homocysteine and genistein: Insights into stroke risk and potential treatment, Metabol. Brain Disease, 32 (2017), 1855–1860. http://doi.org/10.1007/s11011-017-0078-1 doi: 10.1007/s11011-017-0078-1
|
| [66] |
Z. Shi, Y. Guan, Y. R. Huo, S. Liu, M. Zhang, H. Lu, et al., Elevated total homocysteine levels in acute ischemic stroke are associated with long-term mortality, Stroke, 46 (2015), 2419–2425. http://doi.org/10.1161/strokeaha.115.009136 doi: 10.1161/strokeaha.115.009136
|
Figures(8)

Rongxing Qin, Lijuan Huang, Wei Xu, Qingchun Qin, Xiaojun Liang, Xinyu Lai, Xiaoying Huang, Minshan Xie, Li Chen. Identification of disulfidptosis-related genes and analysis of immune infiltration characteristics in ischemic strokes[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18939-18959. doi: 10.3934/mbe.2023838







 DownLoad:
DownLoad: