In some regions of the Americas, domestic dogs are the host for the tick vector Rhipicephalus sanguineus, and spread the tick-borne pathogen Rickettsia rickettsii, which causes Rocky Mountain Spotted Fever (RMSF) in humans. Interventions are carried out against the vector via dog collars and acaricidal wall treatments. This paper investigates the optimal control of acaricidal wall treatments, using a prior model for populations and disease transmission developed for this particular vector, host, and pathogen. It is modified with a death term during questing stages reflecting the cost of control and level of coverage. In the presence of the control, the percentage of dogs and ticks infected with Ri. rickettsii decreases in a short period and remains suppressed for a longer period, including after treatment is discontinued. Risk of RMSF infection declines by 90% during this time. In the absence of re-application, infected tick and dog populations rebound, indicating the eventual need for repeated treatment.
Citation: Maeve L. McCarthy, Dorothy I. Wallace. Optimal control of a tick population with a view to control of Rocky Mountain Spotted Fever[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18916-18938. doi: 10.3934/mbe.2023837
In some regions of the Americas, domestic dogs are the host for the tick vector Rhipicephalus sanguineus, and spread the tick-borne pathogen Rickettsia rickettsii, which causes Rocky Mountain Spotted Fever (RMSF) in humans. Interventions are carried out against the vector via dog collars and acaricidal wall treatments. This paper investigates the optimal control of acaricidal wall treatments, using a prior model for populations and disease transmission developed for this particular vector, host, and pathogen. It is modified with a death term during questing stages reflecting the cost of control and level of coverage. In the presence of the control, the percentage of dogs and ticks infected with Ri. rickettsii decreases in a short period and remains suppressed for a longer period, including after treatment is discontinued. Risk of RMSF infection declines by 90% during this time. In the absence of re-application, infected tick and dog populations rebound, indicating the eventual need for repeated treatment.

| [1] |
M. B. Labruna, M. Gerardi, F. S Krawczak, J. Moraes-Filho, Comparative biology of the tropical and temperate species of Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) under different laboratory conditions, Ticks Tick-borne Diseases, 8 (2017), 146–156. https://doi.org/10.1016/j.ttbdis.2016.10.011 doi: 10.1016/j.ttbdis.2016.10.011

|
| [2] |
M. W. Lineberry, A. N. Grant, K. D. Sundstrom, S. E. Little, K. E. Allen, Diversity and geographic distribution of rickettsial agents identified in brown dog ticks from across the United States, Ticks Tick-borne Diseases, 13 (2022), 102050. https://doi.org/10.1016/j.ttbdis.2022.102050 doi: 10.1016/j.ttbdis.2022.102050
|
| [3] |
R. B. McFee, Tick borne illness-Rocky mountain spotted fever, Disease-a-month DM, 64 (2018), 185–194. https://doi.org/10.1016/j.disamonth.2018.01.006 doi: 10.1016/j.disamonth.2018.01.006
|
| [4] |
K. T. Duncan, K. D. Sundstrom, D. Hunt, M. W. Lineberry, A. Grant, S. E. Little, Survey on the Presence of Equine Tick-Borne Rickettsial Infections in Southcentral United States, J. Equine Veter. Sci., 118 (2022), 104135. https://doi.org/10.1016/j.jevs.2022.104135 doi: 10.1016/j.jevs.2022.104135
|
| [5] |
H. K. Kim, Rickettsia-host-tick interactions: Knowledge advances and gaps, Infect. Immun., 90 (2022), e00621–21. https://doi.org/10.1128/iai.00621-21 doi: 10.1128/iai.00621-21
|
| [6] | L. Backus, J. Foley, C. Chung, S. Virata, O. E. Zazueta, A. López-Pérez, Tick-borne pathogens detected in sheltered dogs during an epidemic of Rocky Mountain spotted fever, a One Health challenge, J. Am. Veter. Med. Assoc., 261 (2023), 375–383. |
| [7] |
Y.-Y. Zhang, Y.-Q. Sun, J.-J. Chen, A.-Y. Teng, T. Wang, H. Li, et al., Mapping the global distribution of spotted fever group rickettsiae: a systematic review with modelling analysis, Lancet Digital Health, 5 (2023), e5–e15. https://doi.org/10.1016/S2589-7500(22)00212-6 doi: 10.1016/S2589-7500(22)00212-6
|
| [8] |
A. M. López-Pérez, A. Chaves, S. Sánchez-Montes, P. Foley, M. Uhart, J. Barrón-Rodríguez, et al., Diversity of rickettsiae in domestic, synanthropic, and sylvatic mammals and their ectoparasites in a spotted fever-epidemic region at the western US-Mexico border, Transbound. Emerg. Diseases, 69 (2022), 609–622. https://doi.org/10.1111/tbed.14027 doi: 10.1111/tbed.14027
|
| [9] | C. M. Ribeiro, J. L. B. de Carvalho, P. A. de Santis Bastos, S. Katagiri, E. Y. Batalha, W. Okano, et al., Prevalence of Rickettsia rickettsii in Ticks: Systematic Review and Meta-analysis, Vector-Borne Zoonot. Diseases, 2021. |
| [10] |
L. S. Blanton, The rickettsioses: A practical update, Infect. Disease Clin., 33 (2019), 213–229. https://doi.org/10.1016/j.idc.2018.10.010 doi: 10.1016/j.idc.2018.10.010
|
| [11] |
G. Álvarez-Hernández, J. F. Roldán, N. S. Milan, R. R. Lash, C. B. Behravesh, C. D. Paddock, Rocky Mountain spotted fever in Mexico: Past, present, and future, Lancet Infect. Diseases, 17 (2017), e189–e196. https://doi.org/10.1016/S1473-3099(17)30173-1 doi: 10.1016/S1473-3099(17)30173-1
|
| [12] |
H. G Koch, M. D. Tuck, Molting and survival of the brown dog tick (Acari: Ixodidae) under different temperatures and humidities, Ann. Entomol. Soc. Am., 79 (1986), 11–14. https://doi.org/10.1093/aesa/79.1.11 doi: 10.1093/aesa/79.1.11
|
| [13] |
J. Gray, F. Dantas-Torres, A. Estrada-Peña, M. Levin, Systematics and ecology of the brown dog tick, Rhipicephalus sanguineus, Ticks Tick-borne Diseases, 4 (2013), 171–180. https://doi.org/10.1016/j.ttbdis.2012.12.003 doi: 10.1016/j.ttbdis.2012.12.003
|
| [14] |
F. Dantas-Torres, Climate change, biodiversity, ticks and tick-borne diseases: the butterfly effect, Int. J. Parasitol. Paras. Wildlife, 4 (2015), 452–461. https://doi.org/10.1016/j.ijppaw.2015.07.001 doi: 10.1016/j.ijppaw.2015.07.001
|
| [15] |
M. S. Pérez, T. P. F. Arroyo, C. S. V. Barrera, C. Sosa-Gutiérrez, J. Torres, K. A. Brown, et al., Predicting the impact of climate change on the distribution of Rhipicephalus sanguineus in the Americas, Sustainability, 15 (2023), 4557. https://doi.org/10.3390/su15054557 doi: 10.3390/su15054557
|
| [16] |
E. O. Jones, J. M. Gruntmeir, S. A. Hamer, S. E. Little, Temperate and tropical lineages of brown dog ticks in North America, Veter. Parasitol. Region. Studies Rep., 7 (2017), 58–61. https://doi.org/10.1016/j.vprsr.2017.01.002 doi: 10.1016/j.vprsr.2017.01.002
|
| [17] |
M. Brophy, M. A Riehle, N. Mastrud, A. Ravenscraft, J. E. Adamson, K. R. Walker, Genetic variation in Rhipicephalus sanguineus sl ticks across Arizona, Int. J. Environ. Res. Public Health, 19 (2022), 4223. https://doi.org/10.3390/ijerph19074223 doi: 10.3390/ijerph19074223
|
| [18] |
S. Sánchez-Montes, B. Salceda-Sánchez, S. E. Bermúdez, G. Aguilar-Tipacamú, G. G. Ballados-González, H. Huerta, et al., Rhipicephalus sanguineus complex in the Americas: systematic, genetic diversity, and geographic insights, Pathogens, 10 (2021), 1118. https://doi.org/10.3390/pathogens10091118 doi: 10.3390/pathogens10091118
|
| [19] |
G. E. Zemtsova, D. A. Apanaskevich, W. K. Reeves, M. Hahn, A. Snellgrove, M. L. Levin, Phylogeography of Rhipicephalus sanguineus sensu lato and its relationships with climatic factors, Exper. Appl. Acarol., 69 (2016), 191–203. https://doi.org/10.1007/s10493-016-0035-4 doi: 10.1007/s10493-016-0035-4
|
| [20] |
A. M. Kjemtrup, K. Padgett, C. D. Paddock, S. Messenger, J. K. Hacker, T. Feiszli, et al., A forty-year review of Rocky Mountain spotted fever cases in California shows clinical and epidemiologic changes, PLoS Neglect. Trop. Diseases, 16 (2022), e0010738. https://doi.org/10.1371/journal.pntd.0010738 doi: 10.1371/journal.pntd.0010738
|
| [21] |
R. Rosenberg, N. P. Lindsey, M. Fischer, C. J. Gregory, A. F. Hinckley, P. S. Mead, et al., Vital signs: Trends in reported vectorborne disease cases United States and Territories, 2004–2016, Morbid. Mortal. Weekly Rep., 67 (2018), 496. https://doi.org/10.15585/mmwr.mm6717e1 doi: 10.15585/mmwr.mm6717e1
|
| [22] |
L. Kidd, Emerging Spotted Fever Rickettsioses in the United States, Veter. Clin. Small Animal Pract., 52 (2022), 1305–1317. https://doi.org/10.1016/j.cvsm.2022.07.003 doi: 10.1016/j.cvsm.2022.07.003
|
| [23] |
A. Bishop, J. Borski, H.-H. Wang, T. G. Donaldson, A. Michalk, A. Montgomery, et al., Increasing incidence of spotted fever group rickettsioses in the United States, 2010–2018, Vector-Borne Zoonot. Diseases, 22 (2022), 491–497. https://doi.org/10.1089/vbz.2022.0021 doi: 10.1089/vbz.2022.0021
|
| [24] |
P. P. VP Diniz, M. J. Beall, K. Omark, R. Chandrashekar, D. A. Daniluk, K. E. Cyr, et al., High prevalence of tick-borne pathogens in dogs from an Indian reservation in northeastern Arizona, Vector-Borne Zoonot Diseases, 10 (2010), 117–123. https://doi.org/10.1089/vbz.2008.0184 doi: 10.1089/vbz.2008.0184
|
| [25] |
A. L Wilson, O. Courtenay, L. A. Kelly-Hope, T. W. Scott, W. Takken, S. J. Torr, et al., The importance of vector control for the control and elimination of vector-borne diseases. PLoS neglected tropical diseases, 14 (2000), e0007831. https://doi.org/10.1371/journal.pntd.0007831 doi: 10.1371/journal.pntd.0007831
|
| [26] |
C. Tourapi, C. Tsioutis, Circular policy: A new approach to vector and vector-borne diseases management in line with the global vector control response (2017–2030), Trop. Med. Infect. Disease, 7 (2022), 125. https://doi.org/10.3390/tropicalmed7070125 doi: 10.3390/tropicalmed7070125
|
| [27] |
F. Dantas-Torres, The brown dog tick, Rhipicephalus sanguineus (Latreille, 1806)(Acari: Ixodidae): From taxonomy to control, Veter. Parasitol., 152 (2008), 173–185. https://doi.org/10.1016/j.vetpar.2007.12.030 doi: 10.1016/j.vetpar.2007.12.030
|
| [28] |
R. W. H. Sargent, Optimal control, J. Comput. Appl. Math., 124 (2000), 361–371. https://doi.org/10.1016/S0377-0427(00)00418-0 doi: 10.1016/S0377-0427(00)00418-0
|
| [29] |
H. D. Gaff, E. Schaefer, S. Lenhart, Use of optimal control models to predict treatment time for managing tick-borne disease, J. Biol. Dynam., 5 (2011), 517–530. https://doi.org/10.1080/17513758.2010.535910 doi: 10.1080/17513758.2010.535910
|
| [30] |
G. Alvarez-Hernandez, A. V. Trejo, V. Ratti, M. Teglas, D. I Wallace, Modeling of Control Efforts against Rhipicephalus sanguineus, the Vector of Rocky Mountain Spotted Fever in Sonora Mexico, Insects, 132 (2022), 263. https://doi.org/10.3390/insects13030263 doi: 10.3390/insects13030263
|
| [31] |
G. Alvarez-Hernandez, N. Drexler, C. D. Paddock, J. D. Licona-Enriquez, J. D. la Mora, A. Straily, et al., Community-based prevention of epidemic Rocky Mountain spotted fever among minority populations in Sonora, Mexico, using a One Health approach, Transact. Royal Soc. Trop. Med. Hyg., 114 (2020), 293–300. https://doi.org/10.1093/trstmh/trz114 doi: 10.1093/trstmh/trz114
|
| [32] |
F. Dantas-Torres, Biology and ecology of the brown dog tick, Rhipicephalus sanguineus, Paras. Vectors, 3 (2010), 1–11. https://doi.org/10.1186/1756-3305-3-26 doi: 10.1186/1756-3305-3-26
|
| [33] |
R. Ravindran, P. K. Hembram, G. S. Kumar, K. G. A. Kumar, C. K. Deepa, A. Varghese, Transovarial transmission of pathogenic protozoa and rickettsial organisms in ticks, Parasitol. Res., 122 (2023), 691–704. https://doi.org/10.1007/s00436-023-07792-9 doi: 10.1007/s00436-023-07792-9
|
| [34] |
S. Busenberg, K. L. Cooke, The population dynamics of two vertically transmitted infections, Theoret. Popul. Biol., 33 (1988), 181–198. https://doi.org/10.1016/0040-5809(88)90012-3 doi: 10.1016/0040-5809(88)90012-3
|
| [35] |
C. L. Wright, H. D. Gaff, D. E. Sonenshine, W. L. Hynes, Experimental vertical transmission of rickettsia parkeri in the gulf coast tick, amblyomma maculatum, Ticks Tick-borne Diseases, 6 (2015), 568–573. https://doi.org/10.1016/j.ttbdis.2015.04.011 doi: 10.1016/j.ttbdis.2015.04.011
|
| [36] | R. K. Miller, A. N. Michel, Ordinary Differential Equations, Academic Press, 1982. |
| [37] | M. H. Potter, H. F. Weinberger. Maximum Principles in Differential Equations, Prentice-Hall, 1967. |
| [38] |
Z. Abbasi, I. Zamani, A. H. A. Mehra, M. Shafieirad, A. Ibeas, Optimal control design of impulsive sqeiar epidemic models with application to covid-19, Chaos Solit. Fract., 139 (2020), 110054. https://doi.org/10.1016/j.chaos.2020.110054 doi: 10.1016/j.chaos.2020.110054
|
| [39] |
H. W. Berhe, O. D. Makinde, Computational modelling and optimal control of measles epidemic in human population, Biosystems, 190 (2020), 104102. https://doi.org/10.1016/j.biosystems.2020.104102 doi: 10.1016/j.biosystems.2020.104102
|
| [40] |
H. S. Rodrigues, M. T. T. Monteiro, D. F. M. Torres, Dynamics of dengue epidemics when using optimal control, Math. Comput. Model., 52 (2010), 1667–1673. https://doi.org/10.1016/j.mcm.2010.06.034 doi: 10.1016/j.mcm.2010.06.034
|
| [41] |
M. A. L. Caetano, T. Yoneyama, Optimal and sub-optimal control in dengue epidemics, Opt. Control Appl. Methods, 22 (20014), 63–73. https://doi.org/10.1002/oca.683 doi: 10.1002/oca.683
|
| [42] | W. H. Fleming, R. W. Rishel, Deterministic and Stochastic Optimal Control, Springer-Verlag, 1975. https://doi.org/10.1007/978-1-4612-6380-7 |
| [43] | The MathWorks Inc, MATLAB version: 9.13.0 (R2023a), 2023. |
| [44] |
W. K. Hackbusch, A numerical method for solving parabolic equations with opposite orientations. Computing, 20 (1978), 229. https://doi.org/10.1007/BF02251947 doi: 10.1007/BF02251947
|
| [45] | S. Lenhart, J.T. Workman. Optimal Control Applied to Biological Models, Chapman & Hall, CRC, 2007. https://doi.org/10.1201/9781420011418 |
| [46] |
F. O Okumu, B. Chipwaza, E. P. Madumla, E. Mbeyela, G. Lingamba, J. Moore, et al., Implications of bio-efficacy and persistence of insecticides when indoor residual spraying and long-lasting insecticide nets are combined for malaria prevention, Malaria J., 11 (2012), 1–13. https://doi.org/10.1186/1475-2875-11-378 doi: 10.1186/1475-2875-11-378
|
| [47] |
K. Gunasekaran, S. S. Sahu, P. Jambulingam, P. K. Das, DDT indoor residual spray, still an effective tool to control Anopheles fluviatilis-transmitted Plasmodium falciparum malaria in India, Trop. Med. Int. Health, 10 (2005), 160–168. https://doi.org/10.1111/j.1365-3156.2004.01369.x doi: 10.1111/j.1365-3156.2004.01369.x
|
| [48] |
N. A. Hamid, S. N. M. Noor, J. Susubi, N. R. Isa, R. M. Rodzay, A. M. B. Effendi, et al., Semi-field evaluation of the bio-efficacy of two different deltamethrin formulations against Aedes species in an outdoor residual spraying study, Heliyon, 6 (2020), e03230. https://doi.org/10.1016/j.heliyon.2020.e03230 doi: 10.1016/j.heliyon.2020.e03230
|
| [49] |
R. N. C. Guedes, K. Beins, D. N. Costa, G. E. Coelho, H. S. da S Bezerra, Patterns of insecticide resistance in Aedes aegypti: Meta-analyses of surveys in Latin America and the Caribbean, Pest Manag. Sci., 76 (2020), 2144–2157. https://doi.org/10.1002/ps.5752 doi: 10.1002/ps.5752
|
| [50] |
D. E. Gorla, R. Vargas Ortiz, S. S. Catalá, Control of rural house infestation by Triatoma infestans in the Bolivian Chaco using a microencapsulated insecticide formulation, Paras. Vectors, 8 (2015), 1–8. https://doi.org/10.1186/s13071-015-0762-0 doi: 10.1186/s13071-015-0762-0
|
| [51] |
I. Amelotti, S. S. Catalá, D. E. Gorla, Experimental evaluation of insecticidal paints against Triatoma infestans (Hemiptera: Reduviidae), under natural climatic conditions, Paras. Vectors, 2 (2009), 1–7. https://doi.org/10.1186/1756-3305-2-30 doi: 10.1186/1756-3305-2-30
|
| [52] |
K. M. Maloney, J. Ancca-Juarez, R. Salazar, K. Borrini-Mayori, M. Niemierko, J. O. Yukich, et al., Comparison of insecticidal paint and deltamethrin against Triatoma infestans (Hemiptera: Reduviidae) feeding and mortality in simulated natural conditions, J. Vector Ecol., 38 (2013), 6–11. https://doi.org/10.1111/j.1948-7134.2013.12003.x doi: 10.1111/j.1948-7134.2013.12003.x
|
| [53] |
A. G. Alarico, N. Romero, L. Hernández, S. Catalá, D. Gorla, Residual effect of a micro-encapsulated formulation of organophosphates and piriproxifen on the mortality of deltamethrin resistant Triatoma infestans populations in rural houses of the Bolivian Chaco region, Memorias do Instituto Oswaldo Cruz, 105 (2010), 752–756. https://doi.org/10.1590/S0074-02762010000600004 doi: 10.1590/S0074-02762010000600004
|
| [54] | J. C. P. Dias, A. Jemmio, About an insecticidal paint for controlling Triatoma infestans, in Bolivia, Rev. Soc. Bras. Med. Trop., 2008. |
| [55] |
B. Mosqueira, S. Duchon, F. Chandre, J.-M. Hougard, P. Carnevale, S. Mas-Coma, Efficacy of an insecticide paint against insecticide-susceptible and resistant mosquitoes-Part 1: Laboratory evaluation, Malaria J., 9 (2010), 1–6. https://doi.org/10.1186/1475-2875-9-340 doi: 10.1186/1475-2875-9-340
|
| [56] |
B. Mosqueira, J. Chabi, F. Chandre, M. Akogbeto, J.-M. Hougard, P. Carnevale, et al., Efficacy of an insecticide paint against malaria vectors and nuisance in West Africa-Part 2: Field evaluation, Malaria J., 9 (2010), 1–7. https://doi.org/10.1186/1475-2875-9-341 doi: 10.1186/1475-2875-9-341
|
| [57] |
B. Mosqueira, J. Chabi, F. Chandre, M. Akogbeto, J.-M. Hougard, P. Carnevale, et al., Proposed use of spatial mortality assessments as part of the pesticide evaluation scheme for vector control, Malaria J., 12 (2013), 1–6. https://doi.org/10.1186/1475-2875-12-366 doi: 10.1186/1475-2875-12-366
|
| [58] |
B. Mosqueira, D. D. Soma, M. Namountougou, S. Poda, A. Diabaté, O. Ali, et al., Pilot study on the combination of an organophosphate-based insecticide paint and pyrethroid-treated long lasting nets against pyrethroid resistant malaria vectors in Burkina Faso, Acta Trop., 148 (2015), 162–169. https://doi.org/10.1016/j.actatropica.2015.04.010 doi: 10.1016/j.actatropica.2015.04.010
|
| [59] |
A. Villegas, J. Castañeda, J. Pruñonosa, R. Arce, G. Álvarez, Evaluation of density reduction of Aedes aegypti mosquitoes and biosecurity of the use of a paint containing propoxur in selected houses in Hermosillo, Sonora, Mexico, Preprints, 2021. https://doi.org/10.20944/preprints202111.0538.v1 doi: 10.20944/preprints202111.0538.v1
|
| [60] | G. Acapovi-Yao, D. Kaba, K. Allou, D. D. Zoh, L. K. Tongué, K. E. N'Goran, Assessment of the efficiency of insecticide paint and impregnated nets on tsetse populations: Preliminary study in forest relics of Abidjan, Côte d'Ivoire, West African J. Appl. Ecol., 22 (2014), 17–25. |
| [61] |
D. Ghosh, A. Alim, M. M. Huda, C. M. Halleux, M. Almahmud, P. L. Olliaro, et al., Comparison of Novel Sandfly Control Interventions: A Pilot Study in Bangladesh, Am. J. Trop. Med. Hyg., 105 (2021), 1786. https://doi.org/10.4269/ajtmh.20-0997 doi: 10.4269/ajtmh.20-0997
|
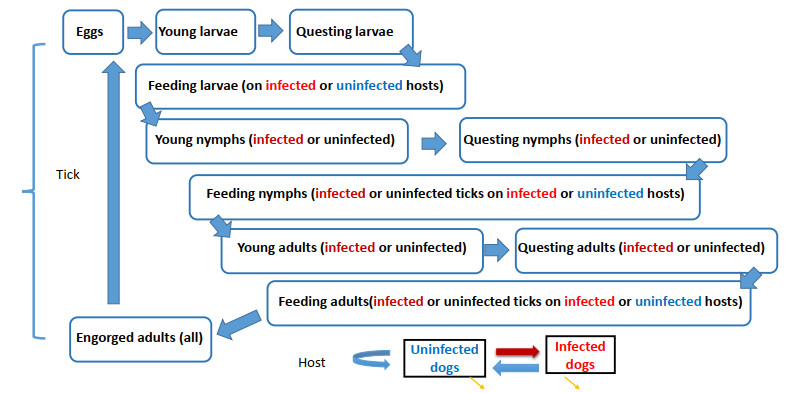
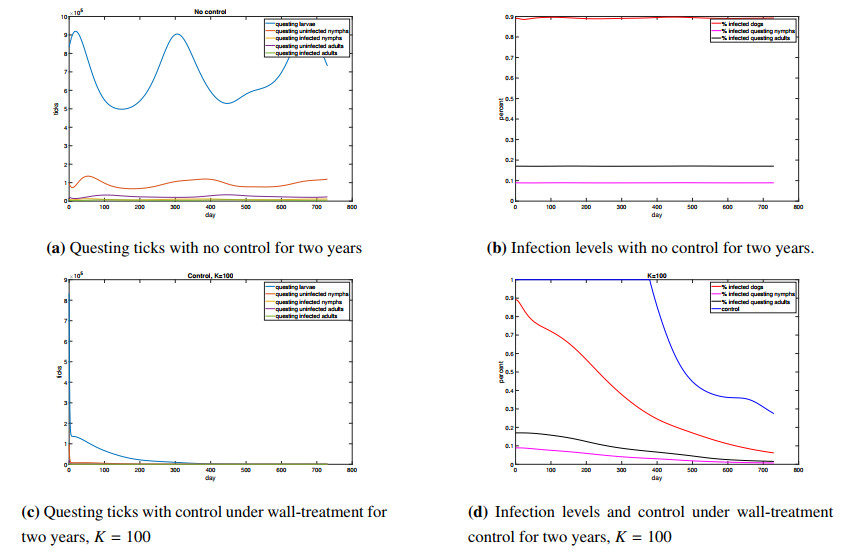
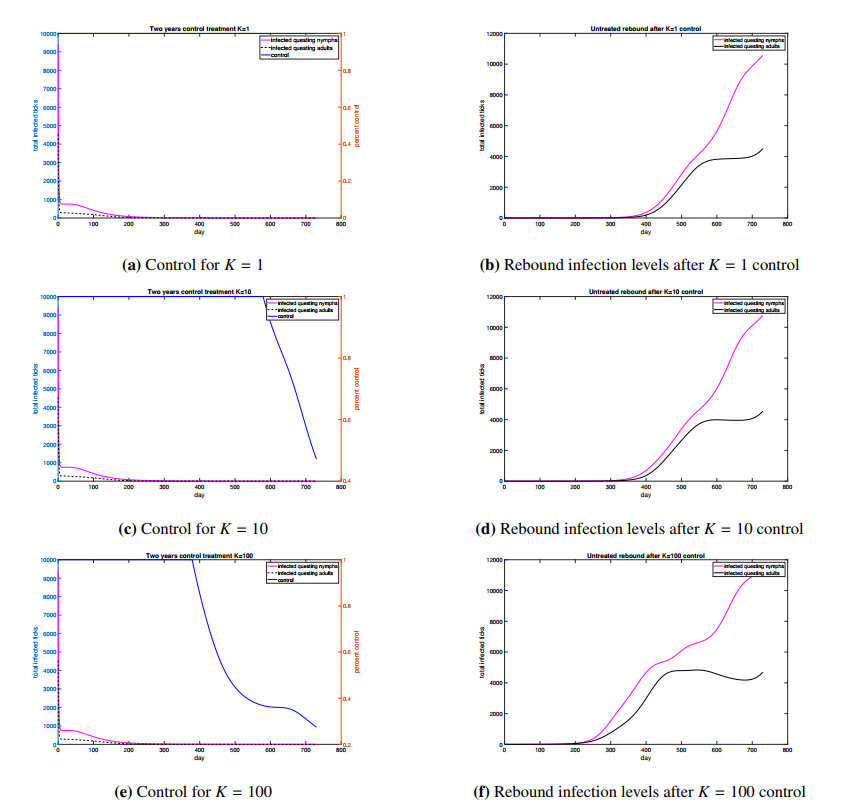
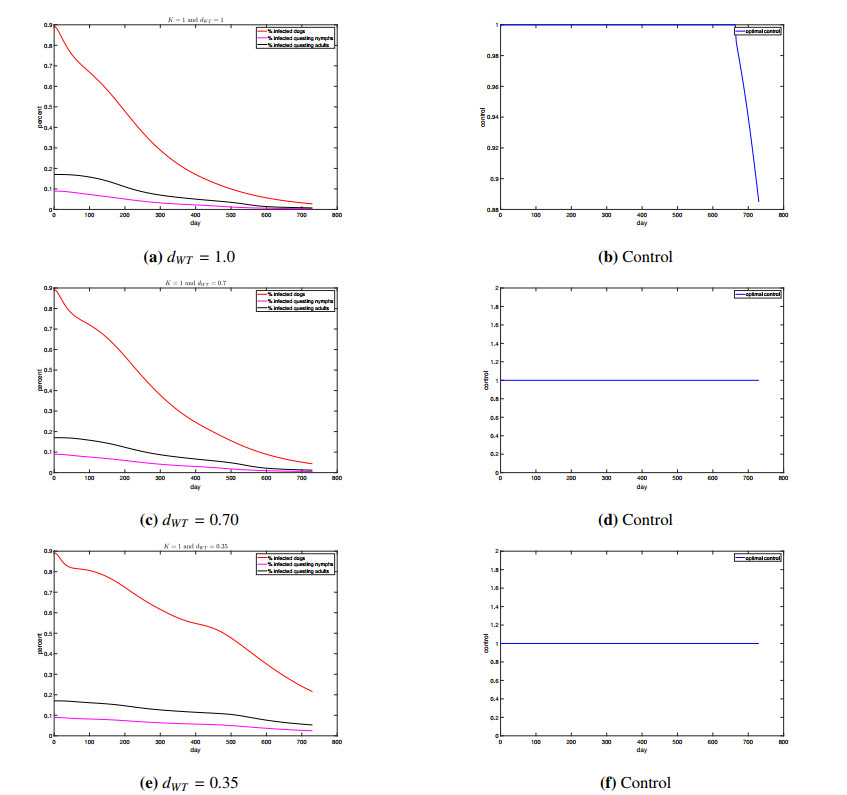
Figures(4) / Tables(1)

Maeve L. McCarthy, Dorothy I. Wallace. Optimal control of a tick population with a view to control of Rocky Mountain Spotted Fever[J]. Mathematical Biosciences and Engineering, 2023, 20(10): 18916-18938. doi: 10.3934/mbe.2023837







 DownLoad:
DownLoad: