Citation: Pei Yu, Xiangyu Wang. Analysis on recurrence behavior in oscillating networks of biologically relevant organic reactions[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5263-5286. doi: 10.3934/mbe.2019263

| [1] | W. Zhang, L. M. Wahl and P. Yu, Modelling and analysis of recurrent autoimmune disease, SIAM J. Appl. Math., 74 (2014), 1998–2025. |
| [2] | H. J. Girschick, C. Zimmer, G. Klaus, et al., Chronic recuurent multifocal osteomyelitis: What is it and how should it be treated? Nat. Clin. Practice Rheumatol., 3 (2007), 733–738. |
| [3] | R. S. Iyer, M. M. Thapa and F. S. Chew, Chronic recurrent multifocal osteomyelitis: Review, Amer. J. Roentgenol., 196 (2011), S87–S91. |
| [4] | D. M. Fergusson, L. J. Horwood and F. T. Shannon, Early solid feeding and recurrent childhood eczema: A 10-year longitudinal study, Pediatrics, 86 (1990), 541–546. |
| [5] | D. D. Munro, Recurrent subacute discoid lupus erythematosus, Proc. Roy. Soc. Med., 56 (1963), 78–79. |
| [6] | T. Vollmer, The natural history of relapses in multiple sclerosis, J. Neurolog. Sci., 256 (2007), s5–s13. |
| [7] | N. Fenichel, Geometri singular perturbation theory, J. Diff. Eqns., 31 (1979) 879–910. |
| [8] | X. F. Chen, P. Yu, M. Han, et al., Canard solutions of 2-D singularly perturbed systems, Chaos Soliton. Fract., 23 (2005), 915–927. |
| [9] | W. Zhang, L. M. Wahl and P. Yu, Conditions for transient viremia in deterministic in-host models: viral blips need no exogenous trigger, SIAM J. Appl. Math., 73 (2013), 853–881. |
| [10] | W. Zhang, L. M. Wahl and P. Yu, Viral blips may not need a trigger: how transient viremia can arise in deterministic in-host models, SIAM Review, 56 (2014), 127–155. |
| [11] | P. Yu, W. Zhang and L.M. Wahl, Dynamical analysis and simulation of a 2-dimensional disease model with convex incidence, Commun. Nonlinear Sci. Numer. Simulat., 37 (2016), 163–192. |
| [12] | N. S. Semenov, J. K. Lewis, A. Alar, et al., Autocatalytic, bistable, oscillatory networks of biolog-ically relevant organic reactions, Nature, 573 (2016), 656–660. |
| [13] | P. Nghe, W. Hordijk, S. A. Kauffman, et al., Prebiotic network evolution:six key parameters, Mol. Biosyst., 11 (2015), 3206–3217. |
| [14] | F. J. Dyson, A model for the origin of life, J. Mol. Evol., 18 (1982), 344–350. |
| [15] | B. H. Patel, C. Percivalle, D. J. Ritson, et al., Common origins ofRNA, protein and lipid precursors in a cyanosulfidic protometabolism, Nat. Chem., 7 (2015), 301–307. |
| [16] | J. J. Tyson, Modeling the cell -division cycle: cdc2 and cyclin interactions, Proc. Natl. Acad. Sci. USA, 88 (1991), 7328–7332. |
| [17] | J. J. Tyson, K. C. Chen, B. Novak, et al., Toggles and blinkers:dynamics of regulatory and signaling pathways in the cell, Curr. Opin. Cell Biol., 15 (2003), 221–231. |
| [18] | J. E. Ferrell, T. Y. C. Tasi and Q. O. Yang, Modeling the cell cycle:why do certain circuits oscillate, Cell, 244 (2011), 874–885. |
| [19] | A. Goldbeter, A model for circadian oscillations in the Drosophila period protein(PER), Proc. R. Soc. Lond., B261 (1995), 319–324. |
| [20] | R. FitzHugh, D. M. Fergusson, L. J. Horwood, et al., Impulses and physiological states in theoret-ical models of nerve membrane, Biophys, J1 (1961), 445–466. |
| [21] | M. T. Laub and W. F. A. Loomis, A molecular network that produces spontaneous oscillations in excitable cells of dictyostelium, Mol. Biol. Cell, 9 (1998), 3521–3532. |
| [22] | A. D. Lander, Pattern, growth, and control, Cell, 144 (2011), 955–969. |
| [23] | B. P. Belousov, Periodicheski deistvuyushchaya reaktsia i ee mechanism [in Russian]. In Sbornik Referatov po Radiatsionni Meditsine, (1958), 145–147. |
| [24] | L. Gyorgyi, T. Turányi and R. J. Field, Mechanistic details of the oscillatory Belousov-Zhabotinskii reaction, J. Phys. Chem., 94 (1990), 7162–7170. |
| [25] | I. R. Epstein and J. A. Pojman, An introduction to Nonlinear Chemical Dynamics: Oscillations, Waves, Patterns, and Chaos, in Chps.2 and 4, Oxford Univ. Press, (1998), 17–47 and 62–83. |
| [26] | I. U. A. Kuznetsov, Elements of Applied Bifurcation Theory, 3rd edition, Ch.3: 77-115, Springer, New York, 2004. |
| [27] | D. Hinrichsen and A. J. Pritchard, Mathematical Systems Theory Ⅰ: Modelling, State Space Anal-ysis, Stability and Robustnes, 2nd edition, Springer-Verlag, New York, 2005. |
| [28] | P. Yu, Closed-form conditions of bifurcation points for general differential equations, Int. J. Bifur-cat. Chaos, 15 (2005), 1467–1483. |
| [29] | M. Han and P. Yu, Normal Forms Melnikov Functions and Bifurcations of Limit Cycles, Springer, London, 2012. |
| [30] | P. Yu, Computation of normal forms via a perturbation technique, J. Sound Vib., 211 (1998), 19–38. |
| [31] | W. Zhang and P. Yu, Hopf and generalized Hopf bifurcations in a recurrent autoimmune disease model, Int. J. Bifurcat. Chaos, 26 (2016), 1650079–1650102. |
| [32] | C. Li, J. Li, Z. Ma, et al., Canard phenomenon for an SIS epidemic model with nonlinear incidence, J. Math. Anal. Appl., 420 (2014), 987–1004. |
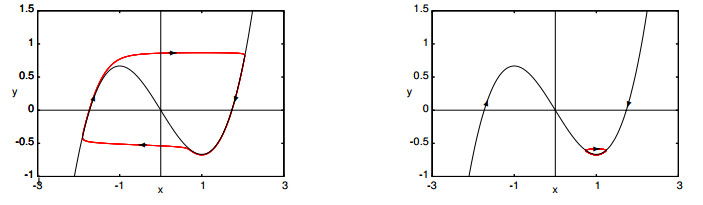
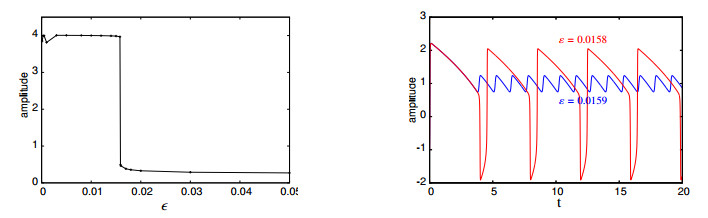
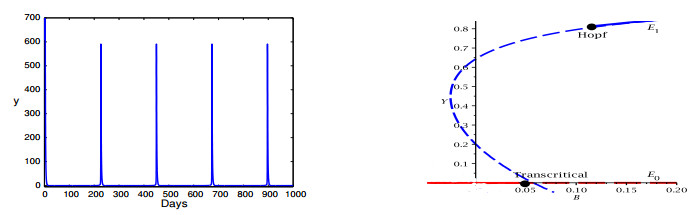
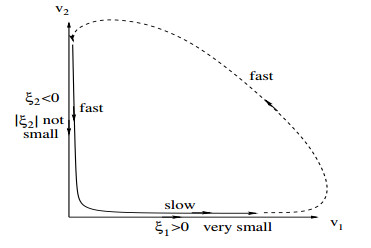
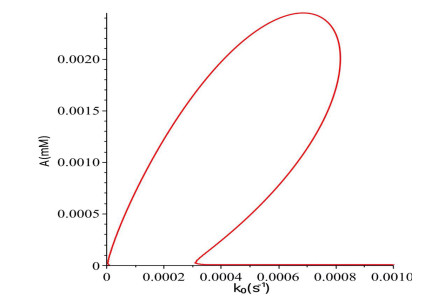
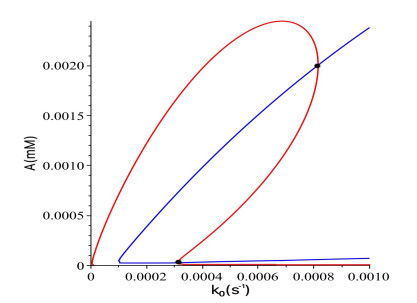
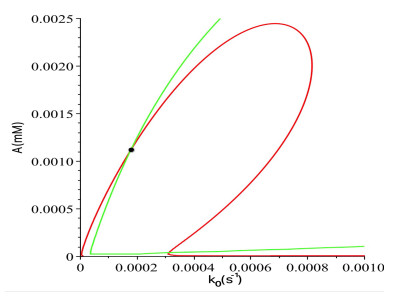
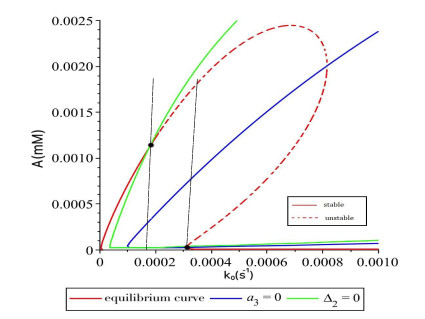
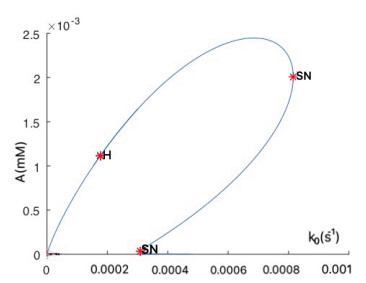
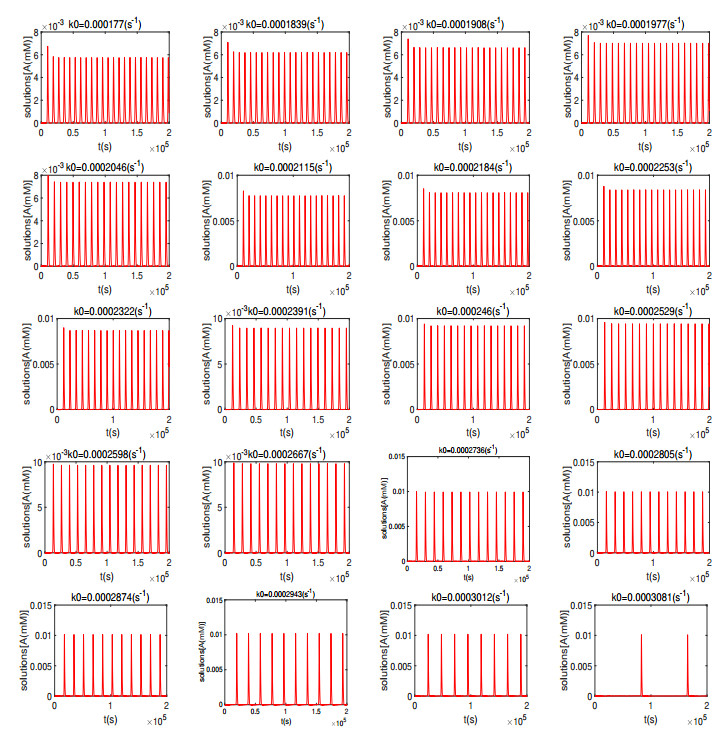
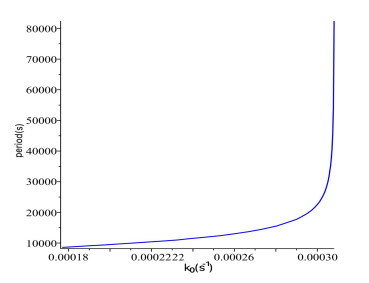
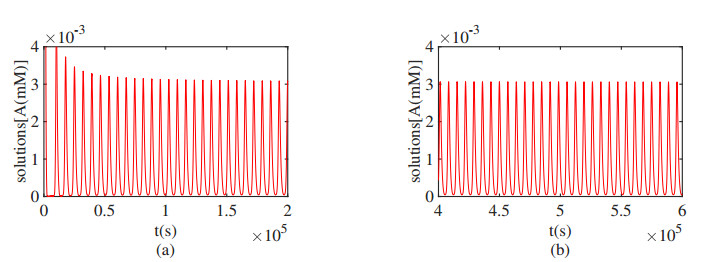
Figures(16)

Pei Yu, Xiangyu Wang. Analysis on recurrence behavior in oscillating networks of biologically relevant organic reactions[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5263-5286. doi: 10.3934/mbe.2019263







 DownLoad:
DownLoad: