Citation: József Z. Farkas, Gary T. Smith, Glenn F. Webb. A dynamic model of CT scans for quantifying doubling time of ground glass opacities using histogram analysis[J]. Mathematical Biosciences and Engineering, 2018, 15(5): 1203-1224. doi: 10.3934/mbe.2018055

| [1] | [ D. Ambrosi,F. Mollica, On the mechanics of a growing tumor, Int. J. Eng. Sci., 40 (2002): 1297-1316. |
| [2] | [ H. Ammari, Mathematical Modeling in Biomedical Imaging 1. Electrical and Ultrasound Tomographies, Anomaly Detection, and Brain Imaging, Springer Science and Business Media, New York, 2009. |
| [3] | [ A. R. A. Anderson,A. M. Weaver,P. T. Cummings,V. Quaranta, Tumor morphology and phenotypic evolution driven by selective pressure from the microenvironment, Cell, 127 (2006): 905-915. |
| [4] | [ F. R. Balkwill,M. Capasso,T. Hagemann, The tumor microenvironment at a glance, J. Cell Sci., 125 (2012): 5591-5596. |
| [5] | [ T. M. Buzug, Computed Tomography, from Photon Statistics to Modern Cone-beam CT, Springer-Verlag, Berlin-Heidelberg-New York, 2008. |
| [6] | [ Á. Calsina,J. Z. Farkas, Positive steady states of nonlinear evolution equations with finite dimensional nonlinearities, SIAM J. Math. Anal., 46 (2014): 1406-1426. |
| [7] | [ R. S. Cantrell,C. Cosner, Diffusive logistic equations with indefinite weights: Population models in disrupted environments Ⅱ, SIAM J. Math. Anal., 22 (1991): 1043-1064. |
| [8] | [ O. Clatz,M. Sermesant,P.-Y. Bondiau,H. Delingette,S. K. Warfield,G. Malandain,N. Ayache, Realistic simulation of the 3-D growth of brain tumors in MR images coupling diffusion with biomechanical deformation, IEEE Trans. Med. Imag., 24 (2005): 1334-1346. |
| [9] | [ F. Cornelis,O. Saut,P. Cumsille,D. Lombardi,A. Iollo,J. Palussiere,T. Colin, In vivo mathematical modeling of tumor growth from imaging data: Soon to come in the future?, Diag. Inter. Imag., 94 (2013): 593-600. |
| [10] | [ H. Enderling,M. A. J. Chaplain, Mathematical modeling of tumor growth and treatment, Curr. Pharm. Des., 20 (2014): 4934-4940. |
| [11] | [ R. A. Gatenby,P. K. Maini,E. T. Gawlinski, Analysis of a tumor as an inverse problem provides a novel theoretical framework for understanding tumor biology and therapy, Appl. Math. Lett., 15 (2002): 339-345. |
| [12] | [ C. I. Henschke,D. F. Yankelevitz,R. Yip,A. P. Reeves,D. Xu,J. P. Smith,D. M. Libby,M. W. Pasmantier,O. S. Miettinen, Lung cancers diagnosed at annual CT screening: Volume doubling times, Radiology, 263 (2012): 578-583. |
| [13] | [ C. I. Henschke,R. Yip,J. P. Smith,A. S. Wolf,R. M. Flores,M. Liang,M. M. Salvatore,Y. Liu,D. M. Xu,D. F. Yankelevitz, CT screening for lung cancer: Part-solid nodules in baseline and annual repeat rounds, Am. J. Roentgenol, 207 (2016): 1176-1184. |
| [14] | [ G. N. Hounsfield, Computed medical imaging, Nobel Lecture, J. Comput. Assist. Tomogr., 4 (1980): 665-674. |
| [15] | [ Y. Kawata,N. Niki,H. Ohmatsu,M. Kusumoto,T. Tsuchida,K. Eguchi,M. Kaneko,N. Moriyama, Quantitative classification based on CT histogram analysis of non-small cell lung cancer: Correlation with histopathological characteristics and recurrence-free survival, Med. Phys., 39 (2012): 988-1000. |
| [16] | [ E. Konukoglu,O. Clatz,B. H. Menze,B. Stieltjes,M-A. Weber,E. Mandonnet,H. Delingette,N. Ayache, Image guided personalization of reaction-diffusion type tumor growth models using modified anisotropic eikonal equations, IEEE Trans. Med. Imag., 29 (2010): 77-95. |
| [17] | [ Y. Kuang, J. D. Nagy and S. E. Eikenberry, Introduction to Mathematical Oncology, Mathematical and Computational Biology Series, Taylor & Francis Group, Boca Raton-London-New York, 2016. |
| [18] | [ J. S. Lowengrub,H. B. Feiboes,F. Jin,Y.-I. Chuang,X. Li,P. Macklin,S. M. Wise,V. Cristini, Nonlinear modelling of cancer: Bridging the gap between cells and tumors, Nonlinearity, 23 (2010): R1-R91. |
| [19] | [ R. H. Martin, Nonlinear Operators and Differential Equations in Banach Spaces, Pure and Applied Mathematics, Wiley-Interscience, John Wiley & Sons, New York-London-Sydney, 1976. |
| [20] | [ D. Morgensztern,K. Politi,R. S. Herbst, EGFR Mutations in non-small-cell lung cancer: Find, divide, and conquer, JAMA Oncol., 1 (2015): 146-148. |
| [21] | [ D. P. Naidich,A. A. Bankier,H. MacMahon,C. M. Schaefer-Prokop,M. Pistolesi,J. M. Goo,P. Macchiarini,J. D. Crapo,C. J. Herold,J. H. Austin,W. D. Travis, Recommendations for the management of subsolid pulmonary nodules detected at CT: A statement from the Fleischner Society, Radiology, 266 (2013): 304-317. |
| [22] | [ National lung screening trial research team, Reduced lung-cancer mortality with low-dose computed tomographic screening, N. Engl. J. Med., 365 (2011), 395–409. |
| [23] | [ J. Prüss, Equilibrium solutions of age-specific population dynamics of several species, J. Math. Biol., 11 (1981): 65-84. |
| [24] | [ R. Rockne, E. C. Alvord, Jr., M. Szeto, S. Gu, G. Chakraborty and K. R. Swanson, Modeling diffusely invading brain tumors: An individualized approach to quantifying glioma evolution and response to therapy, in Selected Topics in Cancer Modeling: Genesis, Evolution, Immune Competition and Therapy, Modeling and Simulation in Science, Engineering and Technology Series, Birkh¨auserBoston, Boston, MA, 2008,207–221. |
| [25] | [ K. R. Swanson,C. Bridge,J. D. Murray,E. C. Alvord Jr., Virtual and real brain tumors: Using mathematical modeling to quantify glioma growth and invasion, J. Neurol. Sci., 216 (2003): 1-10. |
| [26] | [ C. H. Wang,J. K. Rockhill,M. Mrugala,D. L. Peacock,A. Lai,K. Jusenius,J. M. Wardlaw,T. Cloughesy,A. M. Spence,R. Rockne,E. C. Alvord Jr.,K. R. Swanson, Prognostic significance of growth kinetics in newly diagnosed glioblastomas revealed by combining serial imaging with a novel biomathematical model, Can. Res., 69 (2009): 9133-9140. |
| [27] | [ A. Y. Yakovlev, A. V. Zorin and B. I. Grudinko, Computer Simulation in Cell Radiobiology, Lecture Notes in Biomathematics, 74, Springer-Verlag, Berlin-Heidelberg-New York, 1988. |

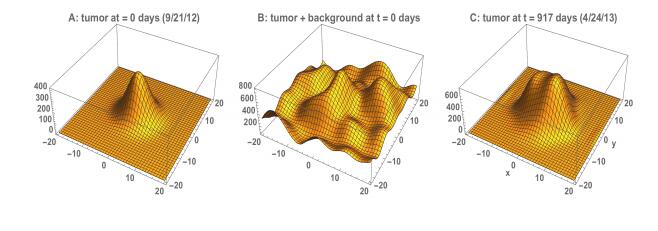
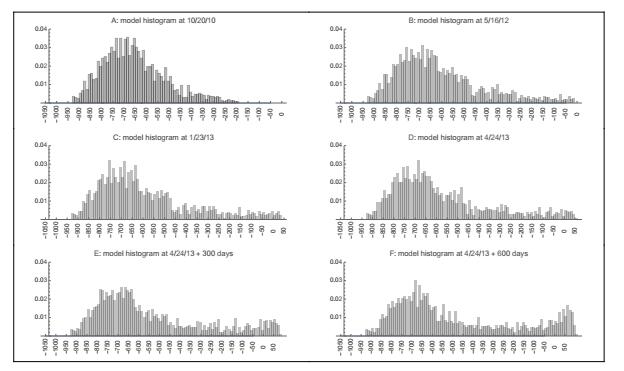
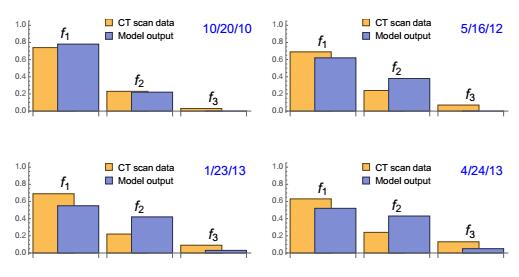
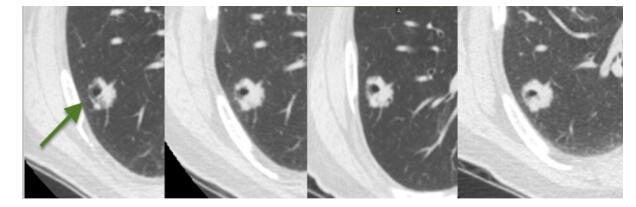
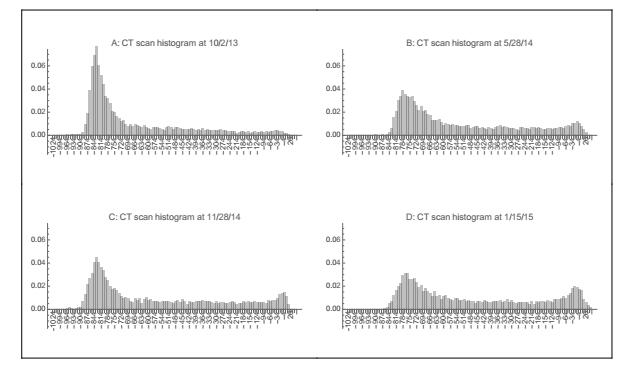
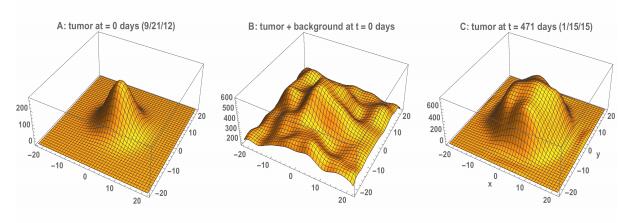
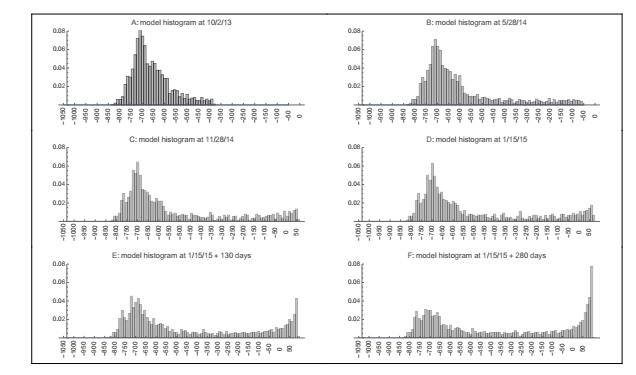
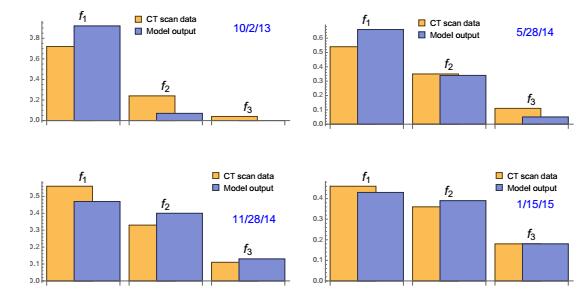
Figures(22) / Tables(3)

József Z. Farkas, Gary T. Smith, Glenn F. Webb. A dynamic model of CT scans for quantifying doubling time of ground glass opacities using histogram analysis[J]. Mathematical Biosciences and Engineering, 2018, 15(5): 1203-1224. doi: 10.3934/mbe.2018055







 DownLoad:
DownLoad: