Citation: Qingwen Hu. A model of regulatory dynamics with threshold-type state-dependent delay[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 863-882. doi: 10.3934/mbe.2018039

| [1] | [ Z. Balanov,Q. Hu,W. Krawcewicz, Global hopf bifurcation of differential equations with threshold type state-dependent delay, J. Differential Equations, 257 (2014): 2622-2670. |
| [2] | [ S. Busenberg,J. M. Mahaffy, Interaction of spatial diffusion and delays in models of genetic control by repression, J. Math. Biol., 22 (1985): 313-333. |
| [3] | [ Y. Chen,J. Wu, Slowly oscillating periodic solutions for a delayed frustrated network of two neurons, J. Math. Anal. Appl., 259 (2001): 188-208. |
| [4] | [ K. Cooke,W. Huang, On the problem of linearization for state-dependent delay differential equations, Proc. Amer. Math. Soc., 124 (1996): 1417-1426. |
| [5] | [ R. D. Driver, A neutral system with state-dependent delay, J. Differential Equations, 54 (1984): 73-86. |
| [6] | [ B. C. Goodwin, Oscillatory behavior in enzymatic control process, Adv. Enzyme Regul., 3 (1965): 425-428. |
| [7] | [ J. Griffith, Mathematics of cellular control processes, ⅰ, negative feedback to one gene, J. Theor. Biol., 20 (1968): 202-208. |
| [8] | [ -, Mathematics of cellular control processes, ⅱ, positive feedback to one gene, J. Theor. Biol, 20 (1968): 209-216. |
| [9] | [ H. Hirata,S. Yoshiura,T. Ohtsuka,Y. Bessho,T. Harada,K. Yoshikawa,R. Kageyama, Oscillatory expression of the bhlh factor hes1 regulated by a negative feedback loop, Science, 298 (2002): 840-843. |
| [10] | [ Q. Hu,W. Krawcewicz,J. Turi, Global stability lobes of a state-dependent model of turning processes, SIAM J. Appl. Math., 72 (2012): 1383-1405. |
| [11] | [ T. Insperger,G. Stépán,J. Turi, State-dependent delay in regenerative turning processes, Nonlinear Dynamics, 47 (2007): 275-283. |
| [12] | [ M. Jensen,K. Sneppen,G. Tiana, Sustained oscillations and time delays in gene expression of protein hes1, FEBS Lett., null (2003): 176-177. |
| [13] | [ J. M. Mahaffy,C. V. Pao, Models of genetic control by repression with time delays and spatial effects, J. Math. Biol., 20 (1984): 39-57. |
| [14] | [ A. H. Nayfeh, Introduction to Perturbation Techniques, John Wiley & Sons, 1981. |
| [15] | [ R. Nussbaum, Limiting profiles for solutions of differential-delay equations, Dynamical Systems V, Lecture Notes in Mathematics, 1822 (2003): 299-342. |
| [16] | [ H. L. Smith, Existence and uniqueness of global solutions for a size-structured model of an insect population with variable instar duration, Rocky Mountain J. Math., 24 (1994): 311-334. |
| [17] | [ H. L. Smith, Oscillations and multiple steady states in a cyclic gene model with repression, J. Math. Biol., 25 (1987): 169-190. |
| [18] | [ P. Smolen,D. A. Baxter,J. H. Byrne, Effects of macromolecular transport and stochastic fluctuations on the dynamics of genetic regulatory systems, Am. J. Physiol., 277 (1999): C777-C790. |
| [19] | [ P. Smolen,D. A. Baxter,J. H. Byrne, Modeling transcriptional control in gene networks-methods, recent results, and future, Bull. Math. Biol., 62 (2000): 247-292. |
| [20] | [ M. Sturrock,A. J. Terry,D. P. Xirodimas,A. M. Thompson,M. A. J. Chaplain, Spatio-temporal modeling of the Hes1 and p53 Mdm2 intracellular signaling pathways, J. Theor. Biol., 273 (2011): 15-31. |
| [21] | [ J. Tyson,H. G. Othmer, The dynamics of feedback control circuits in biochemical pathways, Prog. Theor. Biol., 5 (1978): 2-62. |
| [22] | [ H.-O. Walther, Stable periodic motion of a system using echo for position control, J. Dynam. Differential Equations, 1 (2003): 143-223. |
| [23] | [ -, Smoothness properties of semiflows for differential equations with state-dependent delays, J. Math. Sci., 124 (2004): 5193-5207. |
| [24] | [ A. Wan,X. Zou, Hopf bifurcation analysis for a model of genetic regulatory system with delay, J. Math. Anal. Appl., 356 (2009): 464-476. |
| [25] | [ B. Wu,C. Eliscovich,Y. J. Yoon,R. H. Singer, Translation dynamics of single mRNAs in live cells and neurons, Science, 352 (2016): 1430-1435. |
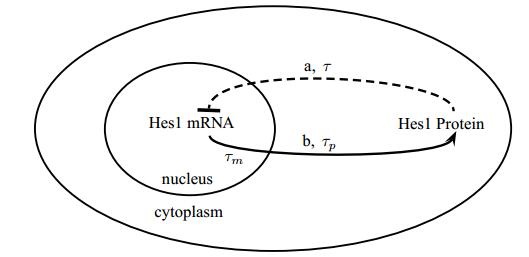
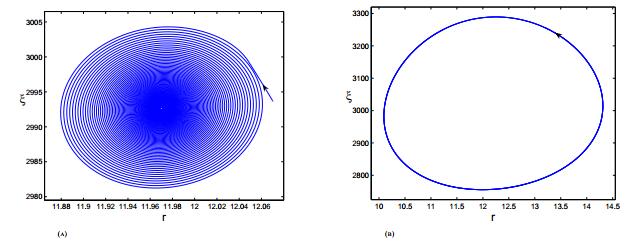
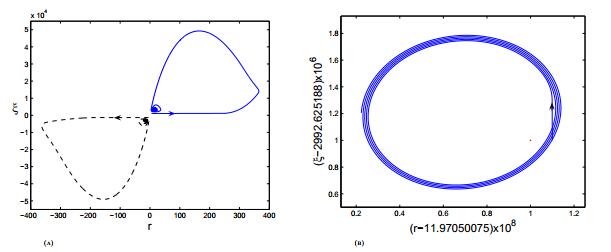
Figures(3)

Qingwen Hu. A model of regulatory dynamics with threshold-type state-dependent delay[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 863-882. doi: 10.3934/mbe.2018039







 DownLoad:
DownLoad: