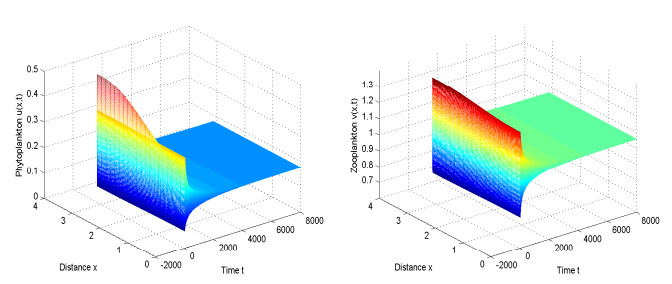
This paper examines a diffusive toxic-producing plankton system with delay. We first show the global attractivity of the positive equilibrium of the system without time-delay. We further consider the effect of delay on asymptotic behavior of the positive equilibrium: when the system undergoes Hopf bifurcation at some points of delay by the normal form and center manifold theory for partial functional differential equations. Global existence of periodic solutions is established by applying the global Hopf bifurcation theory.
Citation: Hong Yang. Global dynamics of a diffusive phytoplankton-zooplankton model with toxic substances effect and delay[J]. Mathematical Biosciences and Engineering, 2022, 19(7): 6712-6730. doi: 10.3934/mbe.2022316
This paper examines a diffusive toxic-producing plankton system with delay. We first show the global attractivity of the positive equilibrium of the system without time-delay. We further consider the effect of delay on asymptotic behavior of the positive equilibrium: when the system undergoes Hopf bifurcation at some points of delay by the normal form and center manifold theory for partial functional differential equations. Global existence of periodic solutions is established by applying the global Hopf bifurcation theory.

| [1] |
S. Chen, H. Yang, J. Wei, Global dynamics of two phytoplankton-zooplankton models with toxic substances effect, J. Appl. Anal. Comput., 9 (2019), 1–14. https://doi.org/10.11948/2156-907X.20180187 doi: 10.11948/2156-907X.20180187

|
| [2] |
J. Chattopadhyay, R. Sarkar, A delay differential equation model on harmful algal blooms in the presence of toxic substances, IMA J. Math. Appl. Med. Biol., 19 (2002), 137–161. https://doi.org/10.1093/imammb/19.2.137 doi: 10.1093/imammb/19.2.137
|
| [3] |
J. Chattopadhayay, R. Sarkar, S. Mandal, Toxin-producing plankton may act as a biological control for planktonic blooms—field study and mathematical modelling, J. Theor. Biol., 215 (2002), 333–344. https://doi.org/10.1006/jtbi.2001.2510 doi: 10.1006/jtbi.2001.2510
|
| [4] |
S. Chaudhuri, J. Chattopadhyay, E. Venturino, Toxic phytoplankton-induced spatiotemporal patterns, J. Biol. Phys., 38 (2012), 331–348. https://doi.org/10.1007/s10867-011-9251-7 doi: 10.1007/s10867-011-9251-7
|
| [5] |
Y. Lv, Y. Pei, S. Gao, C. Li, Harvesting of a phytoplankton-zooplankton model, Nonlinear Anal. Real World Appl., 11 (2010), 3608–3619. https://doi.org/10.1016/j.nonrwa.2010.01.007 doi: 10.1016/j.nonrwa.2010.01.007
|
| [6] |
B. Mukhopadhyay, R. Bhattacharyya, Modelling phytoplankton allelopathy in a nutrient-plankton model with spatial heterogeneity, Ecol.l Modell., 198 (2006), 163–173. https://doi.org/10.1016/j.ecolmodel.2006.04.005 doi: 10.1016/j.ecolmodel.2006.04.005
|
| [7] |
Y. Wang, H. Wang, W. Jiang, Hopf-transcritical bifurcation in toxic phytoplankton-zooplankton model with delay, J. Math. Anal. Appl., 415 (2014), 574–594. https://doi.org/10.1016/j.jmaa.2014.01.081 doi: 10.1016/j.jmaa.2014.01.081
|
| [8] |
J. Zhao, J. Wei, Dynamics in a diffusive plankton system with delay and toxic substances effect, Nonlinear Anal. Real World Appl., 22 (2015), 66–83. https://doi.org/10.1016/j.nonrwa.2014.07.010 doi: 10.1016/j.nonrwa.2014.07.010
|
| [9] |
T. Saha, M. Bandyopadhyay, Dynamical analysis of toxin producing phytoplankton-zooplankton interactions, Nonlinear Anal. Real World Appl., 10 (2009), 314–332. https://doi.org/10.1016/j.nonrwa.2007.09.001 doi: 10.1016/j.nonrwa.2007.09.001
|
| [10] |
J. Zhao, J. P. Tian, J. Wei, Minimal model of plankton systems revisited with spatial diffusion and maturation delay, Bull. Math. Biol., 78 (2016), 381–412. https://doi.org/10.1007/s11538-016-0147-3 doi: 10.1007/s11538-016-0147-3
|
| [11] |
Y. Zhao, Z. Feng, Y. Zheng, X. Cen, Existence of limit cycles and homoclinic bifurcation in a plant-herbivore model with toxin-determined functional response, J. Differe. Equations, 258 (2015), 2847–2872. https://doi.org/10.1016/j.jde.2014.12.029 doi: 10.1016/j.jde.2014.12.029
|
| [12] | R. Peng, J. Shi, M. Wang, On stationary patterns of a reaction-diffusion model with autocatalysis and saturation law, Nonlinearity, 21 (2008), 1471–1488. |
| [13] |
C. Pao, Dynamics of nonlinear parabolic systems with time delays, J. Math. Anal. Appl., 198 (1996), 751–779. https://doi.org/10.1006/jmaa.1996.0111 doi: 10.1006/jmaa.1996.0111
|
| [14] | C. Pao, Convergence of solutions of reaction-diffusion systems with time delays, Nonlinear Anal. Theory Methods Appl., 48 (2002), 349–362. |
| [15] | S. Ruan, J. Wei, On the zeros of transcendental functions with applications to stability of delay differential equations with two delays, Dyn. Contin. Discrete Impulsive Syst. Series A Math. Anal., 10 (2003), 863–874. |
| [16] | B. Hassard, N. Kazarinoff, Y. Wan, Theory and Applications of Hopf Bifurcation, Cambridge University Press, Cambridge, 1981. |
| [17] | J. Wu, Theory and applications of partial functional-differential equations, in Applied Mathematical Sciences, Springer, New York. 1996. |
| [18] | S. Chen, J. Wei, J. Yu, Stationary patterns of a diffusive predator-prey model with Crowley-Martin functional response, Nonlinear Anal. Real World Appl., 39 (2018), 33-57. |
| [19] | D. Gilbarg, N. Trudinger, Elliptic Partial Differential Equation of Second Order, Springer-Verlag, Berlin. 2001. |
| [20] |
T. Kar, K. Chaudhuri, On non-selective harvesting of two competing fish species in the presence of toxicity, Ecol. Modell., 161 (2003), 125–137. https://doi.org/10.1016/S0304-3800(02)00323-X doi: 10.1016/S0304-3800(02)00323-X
|
| [21] |
C. Lin, W. M. Ni, I. Takagi, Large amplitude stationary solutions to a chemotaxis system, J. Differ. Equations, 72 (1988), 1–27. https://doi.org/10.1016/0022-0396(88)90147-7 doi: 10.1016/0022-0396(88)90147-7
|
| [22] | Y. Lou, W. M. Ni, Diffusion, self-diffusion and cross-diffusion, J. Differ. Equations, 131 (1996), 79–131. |
| [23] |
Y. Lv, J. Cao, J. Song, R. Yuan, Y. Pei, Global stability and Hopf-bifurcation in a zooplankton-phytoplankton model, Nonlinear Dyn., 76 (2014), 345–366. https://doi.org/10.1007/s11071-013-1130-2 doi: 10.1007/s11071-013-1130-2
|
| [24] |
A. B. Medvinsky, S. V. Petrovskii, I. A. Tikhonova, H. Malchow, B. L. Li, Spatiotemporal complexity of plankton and fish dynamics, SIAM Rev., 44 (2002), 311–370. https://doi.org/10.1137/S0036144502404442 doi: 10.1137/S0036144502404442
|
| [25] |
W. Ni, M. Wang, Dynamics and patterns of a diffusive Leslie-Gower prey-predator model with strong Allee effect in prey, J. Differ. Equations, 261 (2016), 4244–4274. https://doi.org/10.1016/j.jde.2016.06.022 doi: 10.1016/j.jde.2016.06.022
|
| [26] |
R. Peng, J. Shi, Non-existence of non-constant positive steady states of two Holling type-II predator-prey systems: Strong interaction case, J. Differ. Equations, 247 (2009), 866–886. https://doi.org/10.1016/j.jde.2009.03.008 doi: 10.1016/j.jde.2009.03.008
|
| [27] |
R. Peng, F. Yi, X. Q. Zhao, Spatiotemporal patterns in a reaction-diffusion model with the Degn-Harrison reaction scheme, J. Differ. Equations, 254 (2013), 2465–2498. https://doi.org/10.1016/j.jde.2012.12.009 doi: 10.1016/j.jde.2012.12.009
|
| [28] |
J. P. Shi, X. F. Wang, On the global bifurcation for quasilinear elliptic systems on bounded domains, J. Differ. Equations, 246 (2009), 2788–2812. https://doi.org/10.1016/j.jde.2008.09.009 doi: 10.1016/j.jde.2008.09.009
|
| [29] | J. Smoller, A. Wasserman, Global bifurcation of steady-state solutions, J. Differ. Equations, 39 (1981), 269–290. |
| [30] |
J. Wang, J. Shi, J. Wei, Predator-prey system with strong Allee effect in prey, J. Math. Biol., 62 (2011), 291–331. https://doi.org/10.1007/s00285-010-0332-1 doi: 10.1007/s00285-010-0332-1
|
| [31] |
J. Wang, J. Shi, J. Wei, Dynamics and pattern formation in a diffusive predator-prey system with strong Allee effect in prey, J. Differ. Equations, 251 (2011), 1276–1304. https://doi.org/10.1016/j.jde.2011.03.004 doi: 10.1016/j.jde.2011.03.004
|
| [32] |
J. Wu, Symmetric functional differential equations and neural networks with memory, Trans. Amer. Math. Soc., 350 (1998), 4799–4838. https://doi.org/10.1090/S0002-9947-98-02083-2 doi: 10.1090/S0002-9947-98-02083-2
|
| [33] |
X. Xu, J. Wei, Bifurcation analysis of a spruce budworm model with diffusion and physiological structures, J. Differ. Equations, 262 (2017), 5206–5230. https://doi.org/10.1016/j.jde.2017.01.023 doi: 10.1016/j.jde.2017.01.023
|
| [34] | H. Yang, Analysis of stationary patterns and bifurcations of a diffusive phytoplankton-zooplankton model with toxic substances effect, J. Math. Anal. Appl., Forthecoming, 2023. |
| [35] |
F. Yi, J. Liu, J. Wei, Spatiotemporal pattern formation and multiple bifurcations in a diffusive bimolecular model, Nonlinear Anal. Real World Appl., 11 (2010), 3770–3781. https://doi.org/10.1016/j.nonrwa.2010.02.007 doi: 10.1016/j.nonrwa.2010.02.007
|
| [36] | F. Yi, J. Wei, J. Shi, Bifurcation and spatio-temporal patterns in a diffusive homogenous predator-prey system, J. Differ. Equations, 246 (2009), 1944–1977. |

Figures(2)

Hong Yang. Global dynamics of a diffusive phytoplankton-zooplankton model with toxic substances effect and delay[J]. Mathematical Biosciences and Engineering, 2022, 19(7): 6712-6730. doi: 10.3934/mbe.2022316







 DownLoad:
DownLoad: