Citation: Zhichao Jiang, Xiaohua Bi, Tongqian Zhang, B.G. Sampath Aruna Pradeep. Global Hopf bifurcation of a delayed phytoplankton-zooplankton system considering toxin producing effect and delay dependent coefficient[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3807-3829. doi: 10.3934/mbe.2019188

| [1] | S. Jang, J. Baglama and L. Wu, Dynamics of phytoplankton-zooplankton systems with toxin producing phytoplankton, Appl. Math. Comput., 227 (2014), 717–740. |
| [2] | F. Rao, The complex dynamics of a stochastic toxic-phytoplankton-zooplankton model, Adv. Difference. Equ., 2014 (2014), 22. |
| [3] | A. Sharma, A. Kumar Sharma and K. Agnihotri, Analysis of a toxin producing phytoplanktonzooplankton interaction with Holling IV type scheme and time delay, Nonlinear Dynam., 81 (2015), 13–25. |
| [4] | B. Ghanbari and J. Gómez-Aguilar, Modeling the dynamics of nutrient-phytoplankton-zooplankton system with variable-order fractional derivatives, Chaos Solitons Fractals, 116 (2018), 114–120. |
| [5] | T. Liao, H. Yu and M. Zhao, Dynamics of a delayed phytoplankton-zooplankton system with Crowley-Martin functional response, Adv. Difference. Equ., 2017 (2017), 5. |
| [6] | J. Li, Y. Song and H. Wan, Dynamical analysis of a toxin-producing phytoplankton-zooplankton model with refuge, Math. Biosci. Eng., 14 (2017), 529–557. |
| [7] | Z. Jiang and T. Zhang, Dynamical analysis of a reaction-diffusion phytoplankton-zooplankton system with delay, Chaos Solitons Fractals, 104 (2017), 693–704. |
| [8] | T. Zhang, X. Liu, X. Meng, et al., Spatio-temporal dynamics near the steady state of a planktonic system, Comput. Math. Appl., 75 (2018), 4490–4504. |
| [9] | T. Zhang, Y. Xing, H. Zang, et al., Spatio-temporal patterns in a predator-prey model with hyperbolic mortality, Nonlinear Dynam., 78 (2014), 265–277. |
| [10] | X. Yu, S. Yuan and T. Zhang, The effects of toxin producing phytoplankton and environmental fluctuations on the planktonic blooms, Nonlinear Dynam., 91 (2018), 1653–1668. |
| [11] | Y. Zhao, S. Yuan and T. Zhang, Stochastic periodic solution of a non-autonomous toxic-producing phytoplankton allelopathy model with environmental fluctuation, Commun. Nonlinear Sci. Numer. Simul., 44 (2017), 266– 276. |
| [12] | Y. Zhao, S. Yuan and T. Zhang, The stationary distribution and ergodicity of a stochastic phytoplankton allelopathy model under regime switching, Commun. Nonlinear Sci. Numer. Simul., 37 (2016), 131–142. |
| [13] | Z. Jiang, W. Zhang, J. Zhang, et al., Dynamical analysis of a phytoplankton-zooplankton system with harvesting term and Holling III functional response, Internat. J. Bifur. Chaos Appl. Sci. Engrg., 28 (2018), 1850162. |
| [14] | J. Chattopadhayay, R. Sarkar and S. Mandal, Toxcin-producing plankton may act as a biological control for planktonic blooms-field study and mathematical modeling, J. Theoret. Biol., 215 (2002), 333–344. |
| [15] | J. Dhar, A. Sharma and S. Tegar, The role of delay in digestion of plankton by fish population: A fishery model, J. Nonlinear Sci. Appl., 1 (2008), 13–19. |
| [16] | J. Chattopadhyay, R. Sarkar and A. El Abdllaoui, A delay differential equation model on harmful algal blooms in the presence of toxic substances, IMA J. Math. Appl. Med. Biol., 19 (2002), 137– 161. |
| [17] | T. Zhang, W. Ma and X. Meng, Global dynamics of a delayed chemostat model with harvest by impulsive flocculant input, Adv. Difference. Equ., 2017 (2017), 115. |
| [18] | Y. Tang and L. Zhou, Great time delay in a system with material cycling and delayed biomass growth, IMA J. Appl. Math., 70 (2005), 191–200. |
| [19] | Y. Tang and L. Zhou, Stability switch and Hopf bifurcation for a diffusive prey-predator system with delay, J. Math. Anal. Appl., 334 (2007), 1290–1307. |
| [20] | T. Saha and M. Bandyopadhyay, Dynamical analysis of toxin producing phytoplanktonzooplankton interactions, Nonlinear Anal. Real World Appl., 10 (2009), 314–332. |
| [21] | M. Rehim and M. Imran, Dynamical analysis of a delay model of phytoplankton-zooplankton interaction, Appl. Math. Model., 36 (2012), 638–647. |
| [22] | Y. Wang, W. Jiang and H. Wang, Stability and global Hopf bifurcation in toxic phytoplanktonzooplankton model with delay and selective harvesting, Nonlinear Dynam., 73 (2013), 881–896. |
| [23] | Z. Jiang, W. Ma and D. Li, Dynamical behavior of a delay differential equation system on toxin producing phytoplankton and zooplankton interaction, Japan J. Indust. Appl. Math., 31 (2014), 583–609. |
| [24] | X. Fan, Y. Song and W. Zhao, Modeling cell-to-cell spread of hiv-1 with nonlocal infections, Complexity, 2018 (2018), 2139290. |
| [25] | M. Chi and W. Zhao, Dynamical analysis of two-microorganism and single nutrient stochastic chemostat model with monod-haldane response Function, Complexity, 2019 (2019), 8719067. |
| [26] | N. Gao, Y. Song, X. Wang, et al., Dynamics of a stochastic SIS epidemic model with nonlinear incidence rates, Adv. Difference. Equ., 2019 (2019), 41. |
| [27] | J. Ivlev, Experimental ecology of the feeding of fishes, Yale University Press, New Haven, 1961. |
| [28] | B. Hassard, N. Kazarinoff and Y. Wan, Theory and Application of Hopf Bifurcation, Cambridge University Press, Cambridge, 1981. |
| [29] | Z. Wang, X. Wang, Y. Li, et al., Stability and Hopf bifurcation of fractional-order complexvalued single neuron model with time delay, Internat. J. Bifur. Chaos Appl. Sci. Engrg., 27 (2017), 1750209. |
| [30] | L. Li, Z. Wang, Y. Li, et al., Hopf bifurcation analysis of a complex-valued neural network model with discrete and distributed delays, Appl. Math. Comput, 330 (2018), 152–169. |
| [31] | Z. Jiang and L. Wang, Global Hopf bifurcation for a predator-prey system with three delays, Internat. J. Bifur. Chaos Appl. Sci. Engrg., 27 (2017), 1750108. |
| [32] | Y. Dai, Y. Jia, H. Zhao, et al., Global Hopf bifurcation for three-species ratio-dependent predatorprey system with two delays, Adv. Difference. Equ., 2016 (2016), 13. |
| [33] | J.Wu, Symmetric functional differential equations and neural networks with memory, Trans. Amer. Math. Soc., 35 (1998), 4799–4838. |
| [34] | J. Hale and S. Lunel, Introduction to Functional Differential Equations, Springer-Verlag, New York, 1993. |
| [35] | V. Lakshmikantham and S. Leela, Differential and Integral Inequalities (Theory and Application): Ordinary Differential Equations, Academic Press, New York, 1969. |
| [36] | E. Beretta and Y. Kuang, Geometric stability switch criteria in delay differential systems with delay dependent parameters, SIAM J. Math. Anal., 33 (2002), 1144–1165. |
| [37] | Y. Qu, J. Wei and S. Ruan, Stability and bifurcation analysis in hematopoietic stem cell dynamics with multiple delays, Phys. D, 23 (2010), 2011–2024. |
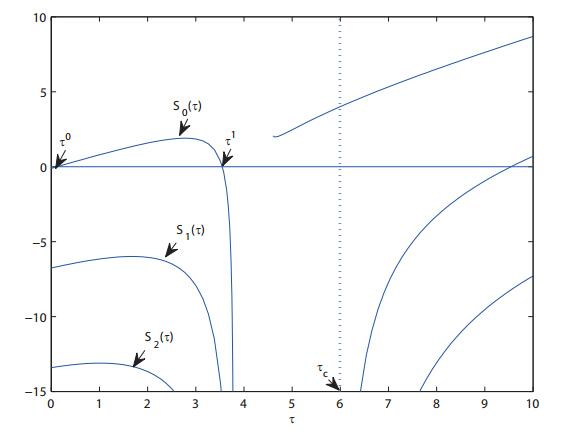
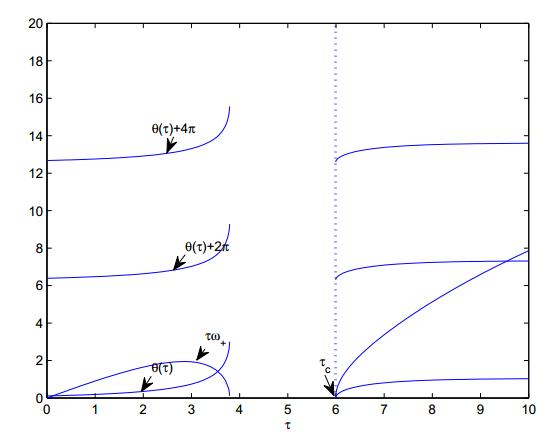
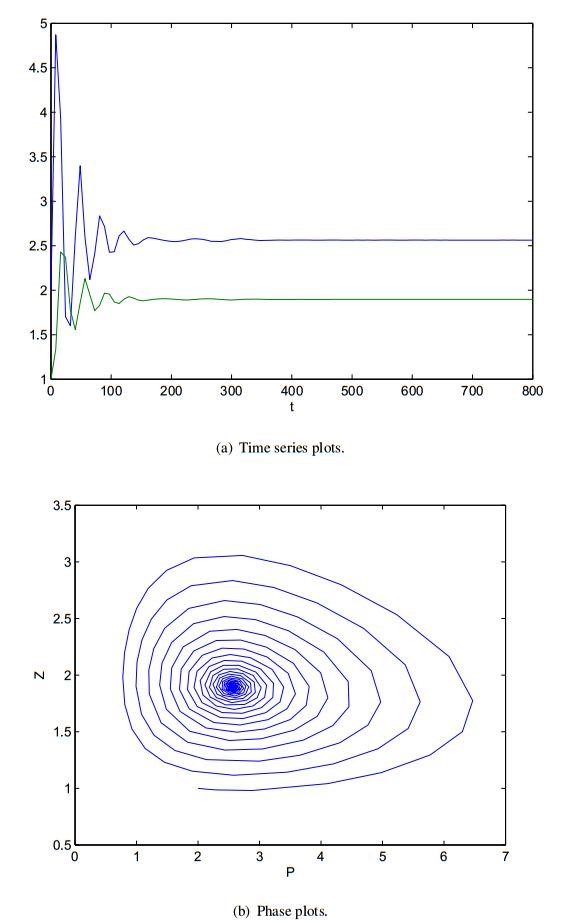
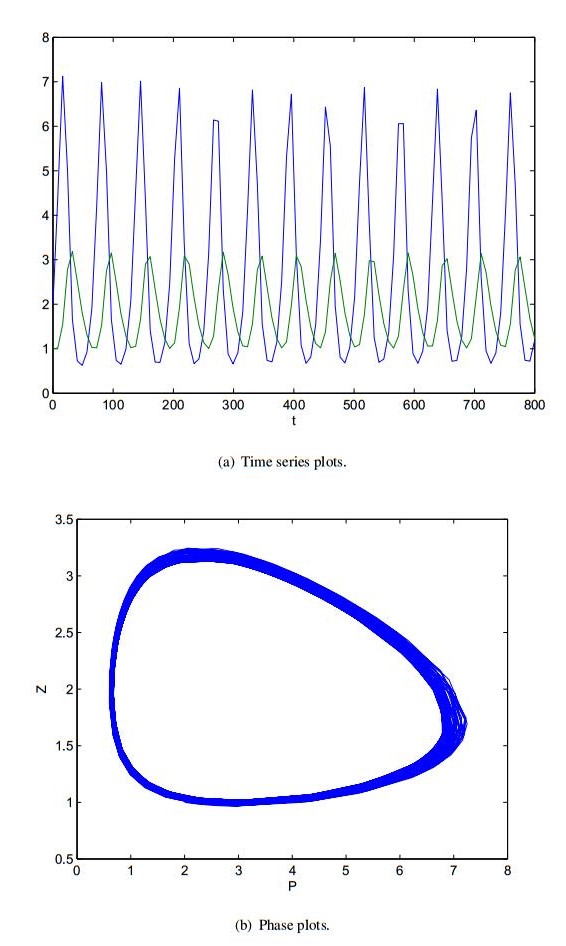
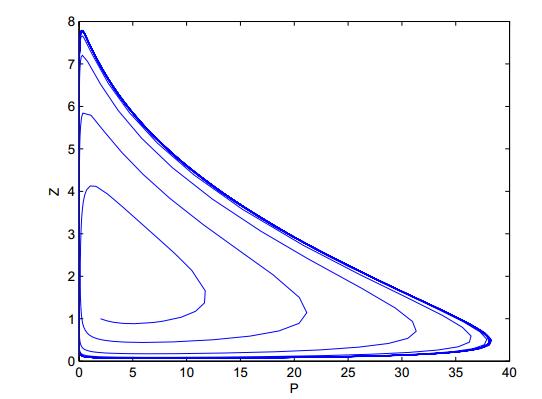
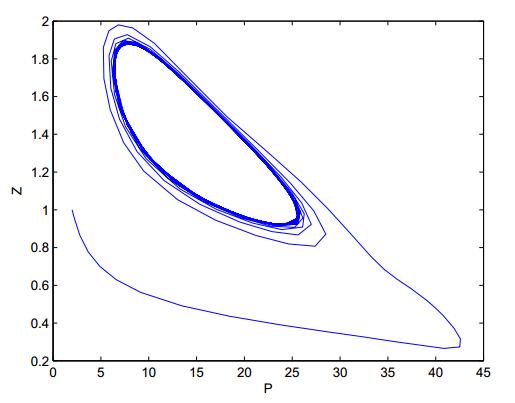
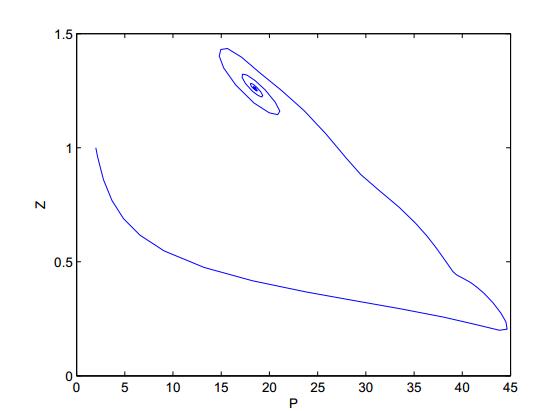
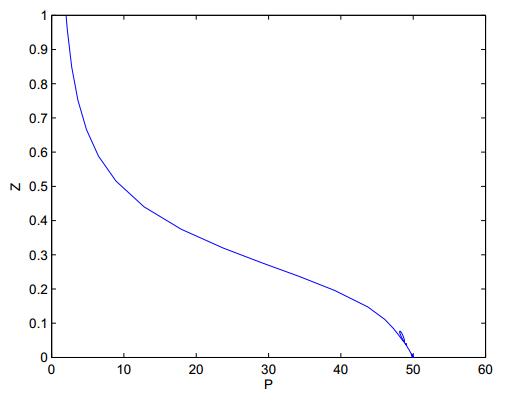
Figures(8)

Zhichao Jiang, Xiaohua Bi, Tongqian Zhang, B.G. Sampath Aruna Pradeep. Global Hopf bifurcation of a delayed phytoplankton-zooplankton system considering toxin producing effect and delay dependent coefficient[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3807-3829. doi: 10.3934/mbe.2019188







 DownLoad:
DownLoad: