Citation: Kalyan Manna, Malay Banerjee. Stability of Hopf-bifurcating limit cycles in a diffusion-driven prey-predator system with Allee effect and time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2411-2446. doi: 10.3934/mbe.2019121

| [1] | G. F. Gause, The Struggle for Existence, Williams and Wilkins: Baltimore, MD, USA, 1935. |
| [2] | A. M. Turing, The chemical basis of morphogenesis, Philos. Trans. R. Soc. Lond., B, Biol. Sci., 237 (1952), 37–72. |
| [3] | S. A. Levin and L. A. Segel, Hypothesis for origin of planktonic patchiness, Nature, 259 (1976), 659. |
| [4] | C. A. Klausmeier, Regular and irregular patterns in semiarid vegetation, Science, 284 (1999), 1826–1828. |
| [5] | H. Malchow, S. V. Petrovskii and E. Venturino, Spatiotemporal Patterns in Ecology and Epidemiology: Theory, Models, and Simulations, Chapman & Hall, London, 2008. |
| [6] | S. V. Petrovskii and H. Malchow, A minimal model of pattern formation in a prey-predator system, Math. Comput. Model., 29 (1999), 49–63. |
| [7] | D. Alonso, F. Bartumeus and J. Catalan, Mutual interference between predators can give rise to Turing spatial patterns, Ecology, 83 (2002), 28–34. |
| [8] | M. Banerjee and S. Petrovskii, Self-organized spatial patterns and chaos in a ratio-dependent predator-prey system, Theor. Ecol., 4 (2011), 37–53. |
| [9] | B. Miao, Persistence and Turing instability in a cross-diffusive predator-prey system with generalist predator, Adv. Differ. Equ., 2018 (2018), 260. |
| [10] | A. B. Medvinsky, B. V. Adamovich and A. Chakraborty, et al., Chaos far away from the edge of chaos: A recurrence quantification analysis of plankton time series, Ecol. Complex., 23 (2015), 61–67. |
| [11] | P. Turchin and S. P. Ellner, Living on the edge of chaos: Population dynamics of Fennoscandian voles, Ecology, 81 (2000), 3099–3116. |
| [12] | W. C. Allee, Animal aggregations: A study in general sociology, University of Chicago Press, Chicago, USA, 1931. |
| [13] | B. Dennis, Allee effect: population growth, critical density, and chance of extinction, Nat. Resour. Model., 3 (1989), 481–538. |
| [14] | P. A. Stephens, W. J. Sutherland and R. P. Freckleton, What is the Allee effect? Oikos, 87 (1999), 185–190. |
| [15] | M. A. Lewis and P. Kareiva, Allee Dynamics and the Spread of Invading Organisms, Theor. Popul. Biol., 43 (1993), 141–158. |
| [16] | E. Odum and G. W. Barrett, Fundamentals of Ecology, Thomson Brooks/Cole, Belmont, CA, 2004. |
| [17] | J. Wang, J. Shi and J. Wei, Predator-prey system with strong Allee effect in prey, J. Math. Biol., 62 (2011), 291–331. |
| [18] | P. Aguirre, E. González-Olivares and E. Sáez, Two limit cycles in a Leslie-Gower predator-prey model with additive Allee effect, Nonlinear Anal. Real World Appl., 10 (2009), 1401–1416. |
| [19] | P. Aguirre, E. González-Olivares and E. Sáez, Three Limit Cycles in a Leslie-Gower Predator-Prey Model with Additive Allee Effect, SIAM J. Appl. Math., 69 (2009), 1244–1262. |
| [20] | K. Manna and M. Banerjee, Stationary, non-stationary and invasive patterns for a prey-predator system with additive Allee effect in prey growth, Ecol. Complex., 36 (2018), 206–217. |
| [21] | Y. Du and J. Shi, Allee effect and bistability in a spatially heterogeneous predator-prey model, Trans. Am. Math. Soc., 359 (2007), 4557–4593. |
| [22] | J. Wang, J. Shi and J. Wei, Dynamics and pattern formation in a diffusive predator-prey system with strong Allee effect in prey, J. Differ. Equ., 251 (2011), 1276–1304. |
| [23] | Y. Cai, W. Wang and J. Wang, Dynamics of a diffusive predator-prey model with additive Allee effect, Int. J. Biomath., 5 (2012), 1250023(11 pages). |
| [24] | Y. Cai, M. Banerjee and Y. Kang, et al., Spatiotemporal complexity in a predator-prey model with weak Allee effects, Math. Biosci. Eng., 11 (2014), 1247–1274. |
| [25] | F. Rao and Y. Kang, The complex dynamics of a diffusive prey-predator model with an Allee effect in prey, Ecol. Complex., 28 (2016), 123–144. |
| [26] | K. Gopalsamy, Stability and Oscillations in Delay Differential Equations of Population Dynamics, Kluwer Academic, Dordrecht, 1992. |
| [27] | Y. Kuang, Delay Differential Equations with Applications in Population Dynamics, Academic Press, New York, 1993. |
| [28] | R. M. May, Time-delay versus stability in population models with two and three trophic levels, Ecology, 54 (1973), 315–325. |
| [29] | S. Ruan, On nonlinear dynamics of predator-prey models with discrete delay, Math. Model. Nat. Pheno., 4 (2009), 140–188. |
| [30] | P. J. Pal, T. Saha and M. Sen, et al., A delayed predator-prey model with strong Allee effect in prey population growth, Nonlinear Dyn., 68 (2012), 23–42. |
| [31] | S. Roy Choudhury, Turing instability in competition models with delay I: Linear theory, SIAM J. Appl. Math., 54 (1994), 1425–1450. |
| [32] | S. Roy Choudhury, Analysis of spatial structure in a predator-prey model with delay II: Nonlinear theory, SIAM J. Appl. Math., 54 (1994), 1451–1467. |
| [33] | K. P. Hadeler and S. Ruan, Interaction of diffusion and delay, Discrete Contin. Dyn. Syst. Ser. B, 8 (2007), 95–105. |
| [34] | S. Sen, P. Ghosh and S. S. Riaz, et al., Time-delay-induced instabilities in reaction-diffusion systems, Phys. Rev. E, 80 (2009), 046212. |
| [35] | S. Chen and J. Shi, Stability and Hopf bifurcation in a diffusive logistic population model with nonlcal delay effect, J. Differ. Equ., 253 (2012), 3440–3470. |
| [36] | C. Tian and L. Zhang, Delay-driven irregular spatiotemporal patterns in a plankton system, Phys. Rev. E, 88 (2013), 012713. |
| [37] | M. Banerjee and L. Zhang, Influence of discrete delay on pattern formation in a ratio-dependent prey-predator model, Chaos Solitons Fractals, 67 (2014), 73–81. |
| [38] | M. Banerjee and L. Zhang, Time delay can enhance spatio-temporal chaos in a prey-predator model, Ecol. Complex., 27 (2016), 17–28. |
| [39] | Y. Song, Y. Peng and X. Zou, Persistence, Stability and Hopf bifurcation in a diffusive ratiodependent predator-prey model with delay, Int. J. Bifurc. Chaos, 24 (2014), 1450093. |
| [40] | S. Chen, J. Shi and J. Wei, Global stability and Hopf bifurcation in a delayed diffusive Leslie- Gower predator-prey system, Int. J. Bifurc. Chaos, 22 (2012), 1250061. |
| [41] | H. Fang, L. Hu and Y.Wu, Delay-induced Hopf bifurcation in a diffusive Holling-Tanner predatorprey model with ratio-dependent response and Smith growth, Adv. Differ. Equ., 2018 (2018), 285. |
| [42] | T. Faria, Stability and bifurcation for a delayed predator-prey model and the effect of diffusion, J. Math. Anal. Appl., 254 (2001), 433–463. |
| [43] | Z. Ge and Y. He, Diffusion effect and stability analysis of a predator-prey system descried by a delayed reaction-diffusion equations, J. Math. Anal. Appl., 339 (2008), 1432–1450. |
| [44] | G. P. Hu andW. T. Li, Hopf bifurcation analysis for a delayed predator-prey system with diffusion effects, Nonlinear Anal. Real World Appl., 11 (2010), 819–826. |
| [45] | J. Li, Z. Jin and G. Q. Sun, Periodic solutions of a spatiotemporal predator-prey system with additional food, Chaos Solitons Fractals, 91 (2016), 350–359. |
| [46] | F. Rao, C. Castillo-Chavez and Y. Kang, Dynamics of a diffusion reaction prey-predator model with delay in prey: Effects of delay and spatial components, J. Math. Anal. Appl., 461 (2018), 1177–1214. |
| [47] | S. Ruan and X. Q. Zhao, Persistence and Extinction in two species reaction-diffusion systems with delays, J. Differ. Equ., 156 (1999), 71–92. |
| [48] | X. Tang and Y. Song, Stability, Hopf bifurcations and spatial patterns in a delayed diffusive predator-prey model with herd behavior, Appl. Math. Comput., 254 (2015), 375–391. |
| [49] | B. Wang, A. L. Wang and Y. J. Liu, et al., Analysis of a spatial predator-prey model with delay, Nonlinear Dyn., 62 (2010), 601–608. |
| [50] | C. Xu and S. Yuan, Spatial periodic solutions in a delayed diffusive predator-prey model with herd behavior, Int. J. Bifurc. Chaos, 25 (2015), 1550155. |
| [51] | X. P. Yan, Stability and Hopf bifurcation for a delayed prey-predator system with diffusion effects, Appl. Math. Comput., 192 (2007), 552–566. |
| [52] | R. Yang, H. Ren and X. Cheng, A diffusive predator-prey system with prey refuge and gestation delay, Adv. Differ. Equ., 2017 (2017), 158. |
| [53] | J. F. Zhang, W. T. Li and X. P. Yan, Bifurcation and spatiotemporal patterns in a homogeneous diffusion-competition system with delays, Int. J. Biomath., 5 (2012), 1250049. |
| [54] | J. Zhao and J. Wei, Persistence, Turing instability and Hopf bifurcation in a diffusive plankton system with delay and quadratic closure, Int. J. Bifurc. Chaos, 26 (2016), 1650047. |
| [55] | W. Zuo, Global stability and Hopf bifurcations of a Beddington-DeAngelis type predator-prey system with diffusion and delays, Appl. Math. Comput., 223 (2013), 423–435. |
| [56] | W. Zuo and J. Wei, Stability and Hopf bifurcation in a diffusive predator-prey system with delay effect, Nonlinear Anal. Real World Appl., 12 (2011), 1998–2011. |
| [57] | M. Jankovic, S. Petrovskii and M. Banerjee, Delay driven spatiotemporal chaos in single species population dynamics models, Theor. Popul. Biol., 110 (2016), 51–62. |
| [58] | Z. P. Ma, W. T. Li and X. P. Yan, Stability and Hopf bifurcation for a three-species food chain model with time delay and spatial diffusion, Appl. Math. Comput., 219 (2012), 2713–2731. |
| [59] | C. Tian and L. Zhang, Hopf bifurcation analysis in a diffusive food-chain model with time delay, Comput. Math. Appl., 66 (2013), 2139–2153. |
| [60] | G. X. Yang and J. Xu, Stability and Hopf bifurcation for a three-species reaction-diffusion predator-prey system with two delays, Int. J. Bifurc. Chaos, 23 (2013), 1350194. |
| [61] | R. Arditi, L. F. Bersier and R. P. Rohr, The perfect mixing paradox and the logistic equation: Verhulst vs. Lotka, Ecosphere, 7 (2016), e01599. |
| [62] | J. P. Gabriel, F. Saucy and L. F. Bersier, Paradoxes in the logistic equation? Ecol. Model., 185 (2005), 147–151. |
| [63] | L. R. Ginzburg, Evolutionary consequences of basic growth equations, TREE, 7 (1992), 133. |
| [64] | J. Mallet, The struggle for existence: How the notion of carrying capacity, K, obscures the links between demography, Darwinian evolution, and speciation, Evol. Ecol. Res., 14 (2012), 627–665. |
| [65] | W. E. Fitzgibbon, Semilinear functional differential equations in Banach space, J. Differ. Equ., 29 (1978), 1–14. |
| [66] | R. H. Martin and H. L. Smith, Abstract functional-differential equations and reaction-diffusion systems, Trans. Am. Math. Soc., 321 (1990), 1–44. |
| [67] | R. H. Martin and H. L. Smith, Reaction-diffusion systems with time delays: Monotonicity, invariance, comparison and convergence, J. reine angew. Math., 413 (1991), 1–35. |
| [68] | C. C. Travis and G. F. Webb, Existence and stability for partial functional differential equations, Trans. Am. Math. Soc., 200 (1974), 395–418. |
| [69] | J. Wu, Theory and Applications of Partial Functional Differential Equations, Springer-Verlag, New York, 1996. |
| [70] | Z. Wu, J. Yin and C. Wang, Elliptic and Parabolic Equations, World Scientific, 2006. |
| [71] | M. H. Protter and H. F.Weinberger, Maximum Principles in Differential Equations, Prentice Hall, Englewood Cliffs, 1967. |
| [72] | M. Sen, M. Banerjee and E. Venturino, A model for biological control in agriculture, Math. Comput. Simul., 87 (2013), 30–44. |
| [73] | X. Song and L. Chen, Optimal harvesting and stability for a two-species competitive system with stage structure, Math. Biosci., 170 (2001), 173–186. |
| [74] | D. Henry, Geometric Theory of Semilinear Parabolic Equations, Lecture Notes in Mathematics, Vol. 840, Springer-Verlag, Berlin, New York, 1981. |
| [75] | B. Hassard, N. Kazarinoff and Y. Wan, Theory and applications of Hopf bifurcation, Cambridge University Press, Cambridge, 1981. |
| [76] | H. I. Freedman and V. S. H. Rao, The trade-off between mutual interference and time lags in predator-prey system, Bull. Math. Biol., 45 (1983), 991–1004. |
| [77] | T. Faria, Normal forms and Hopf bifurcation for partial differential equations with delays, Trans. Am. Math. Soc., 352 (2000), 2217–2238. |
| [78] | A. Morozov, S. Petrovskii and B. L. Li, Spatiotemporal complexity of patchy invasion in a predator-prey system with the Allee effect, J. Theor. Biol., 238 (2006), 18–35. |
| [79] | N. Mukherjee, S. Ghorai and M. Banerjee, Effects of density dependent cross-diffusion on the chaotic patterns in a ratio-dependent prey-predator model, Ecol. Complex., 36 (2018), 276–289. |
| [80] | M. Pascual, Diffusion-induced chaos in a spatial predator-prey system, Proc. R. Soc. Lond., B, Biol. Sci., 251 (1993), 1–7. |
| [81] | A. Wolf, J. B. Swift and H. L. Swinney, et al., Determining Lyapunov exponents from a time series, Physica D, 16 (1985), 285–317. |
| [82] | M. Baurmann, T. Gross and U. Feudel, Instabilities in spatially extended predator-prey systems: Spatio-temporal paterns in the neighborhood of Turing-Hopf bifurcations, J. Theor. Biol., 245 (2007), 220–229. |
| [83] | S. V. Petrovskii and H. Malchow, Wave of chaos: New mechanism of pattern formation in spatiotemporal population dynamics, Theor. Popul. Biol., 59 (2001), 157–174. |
| [84] | E. Ranta, V. Kaitala and J. Lindström, et al., Synchrony in population dynamics, Proc. R. Soc. Lond., B, Biol. Sci., 262 (1995), 113–118. |
| [85] | M. Rietkerk and J. van de Koppel, Regular pattern formation in real ecosystems, Trends Ecol. Evol., 23 (2008), 169–175. |
| [86] | P. Kareiva, A. Mullen and R. Southwood, Population dynamics in spatially complex environments: Theory and data (and discussion), Philos. Trans. R. Soc. Lond., B, Biol. Sci., 330 (1990), 175–190. |
| [87] | T. M. Powell, P. J. Richerson and T. M. Dillon, et al., Spatial scales of current speed and phytoplankton biomass fluctuations in Lake Tahoe, Science, 189 (1975), 1088–1090. |
| [88] | A. A. Sharov, A. M. Liebhold and E. A. Roberts, Correlation of counts of gypsy moths (Lepidoptera: Lymantriidae) in pheromone traps with landscape characteristics, Forest Sci., 43 (1997), 483–490. |
| [89] | A. Okubo, Diffusion and Ecological Problems: Mathematical Models, Springer-Verlag, Berlin, 1980. |
| [90] | A. Okubo and S. A. Levin, Diffusion and Ecological Problems: Modern Perspectives, Springer, Berlin, 2001. |
| [91] | L. A. Segel and J. L. Jackson, Dissipative structure: An explanation and an ecological example, J. Theor. Biol., 37 (1972), 545–559. |
| [92] | Y. V. Tyutyunov, L. I. Titova and I. N. Senina, Prey-taxis destabilizes homogeneous stationary state in spatial Gause-Kolmogorov-type model for predator-prey system, Ecol. Complex., 31 (2017), 170–180. |
| [93] | J. D. Murray, Mathematical Biology, Springer, Heidelberg, 1989. |
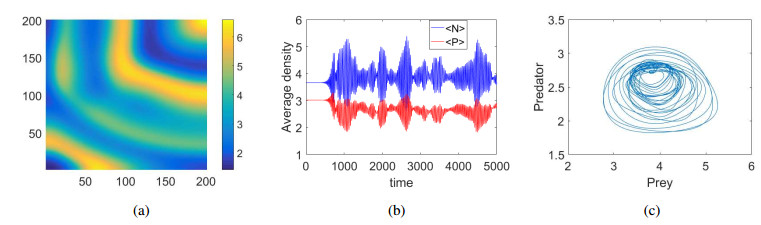
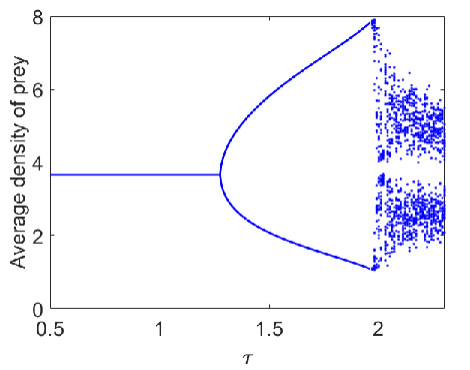
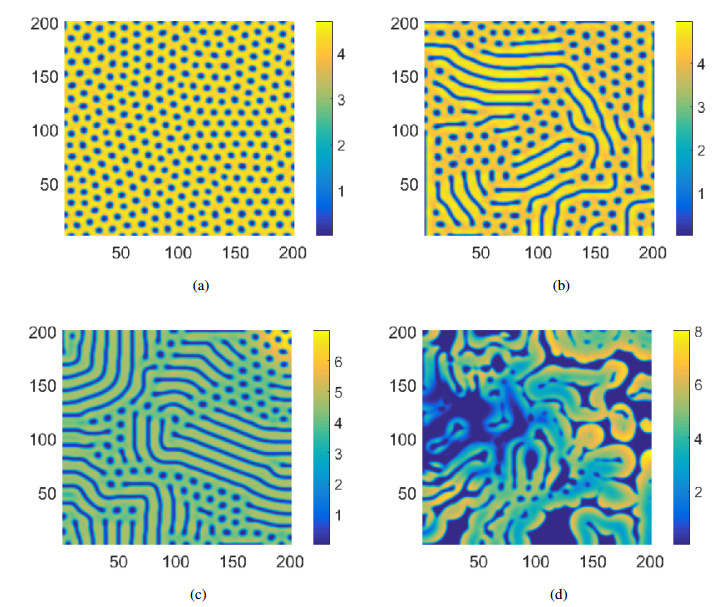
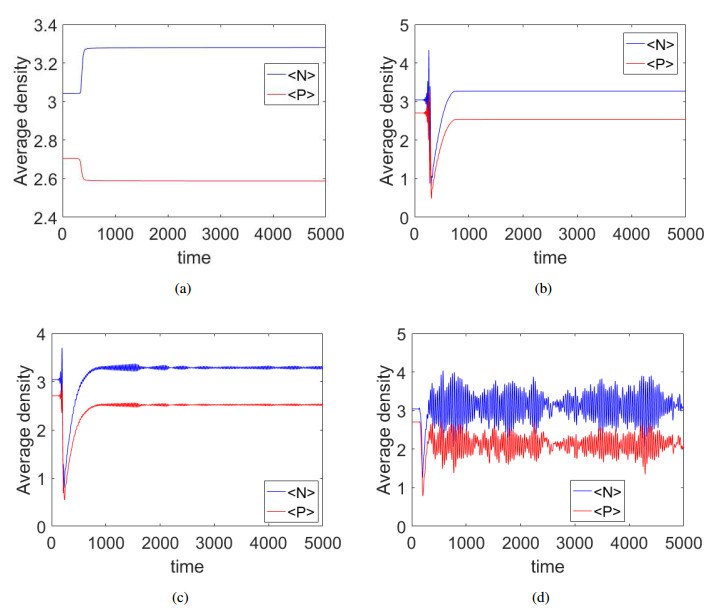
Figures(9)

Kalyan Manna, Malay Banerjee. Stability of Hopf-bifurcating limit cycles in a diffusion-driven prey-predator system with Allee effect and time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2411-2446. doi: 10.3934/mbe.2019121







 DownLoad:
DownLoad: