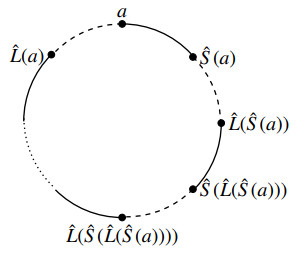
Many structures in science, engineering, and art can be viewed as curves in 3-space. The entanglement of these curves plays a crucial role in determining the functionality and physical properties of materials. Many concepts in knot theory provide theoretical tools to explore the complexity and entanglement of curves in 3-space. However, classical knot theory focuses on global topological properties and lacks the consideration of local structural information, which is critical in practical applications. In this work, two localized models based on the Jones polynomial were proposed, namely, the multi-scale Jones polynomial and the persistent Jones polynomial. The stability of these models, especially the insensitivity of the multi-scale and persistent Jones polynomial models to small perturbations in curve collections, was analyzed, thus ensuring their robustness for real-world applications.
Citation: Ruzhi Song, Fengling Li, Jie Wu, Fengchun Lei, Guo-Wei Wei. Multi-scale Jones polynomial and persistent Jones polynomial for knot data analysis[J]. AIMS Mathematics, 2025, 10(1): 1463-1487. doi: 10.3934/math.2025068
Many structures in science, engineering, and art can be viewed as curves in 3-space. The entanglement of these curves plays a crucial role in determining the functionality and physical properties of materials. Many concepts in knot theory provide theoretical tools to explore the complexity and entanglement of curves in 3-space. However, classical knot theory focuses on global topological properties and lacks the consideration of local structural information, which is critical in practical applications. In this work, two localized models based on the Jones polynomial were proposed, namely, the multi-scale Jones polynomial and the persistent Jones polynomial. The stability of these models, especially the insensitivity of the multi-scale and persistent Jones polynomial models to small perturbations in curve collections, was analyzed, thus ensuring their robustness for real-world applications.

| [1] | R. H. Crowell, R. H. Fox, Introduction to knot theory, New York: Springer, 1963. https://dx.doi.org/10.1007/978-1-4612-9935-6 |
| [2] |
J. W. Alexander, Topological invariants of knots and links, Trans. Amer. Math. Soc., 30 (1928), 275–306. https://dx.doi.org/10.1090/S0002-9947-1928-1501429-1 doi: 10.1090/S0002-9947-1928-1501429-1

|
| [3] |
V. F. R. Jones, A polynomial invariant for knots via von Neumann algebras, Bull. Amer. Math. Soc., 12 (1985), 103–111. https://dx.doi.org/10.1090/s0273-0979-1985-15304-2 doi: 10.1090/s0273-0979-1985-15304-2
|
| [4] | C. Manolescu, An introduction to knot Floer homology, 2014, arXiv: 1401.7107. |
| [5] |
M. Khovanov, A categorification of the Jones polynomial, Duke Math. J., 101 (2000), 359–426. https://dx.doi.org/10.1215/S0012-7094-00-10131-7 doi: 10.1215/S0012-7094-00-10131-7
|
| [6] | T. Ohtsuki, Quantum invariants: A study of knots, 3-manifolds, and their sets, Singapore: World Scientific, 2001. https://dx.doi.org/10.1142/4746 |
| [7] | C. Z. Liang, K. Mislow, Knots in proteins, J. Am. Chem. Soc., 116 (1994), 11189–11190. https://dx.doi.org/10.1021/ja00103a057 |
| [8] | D. W. Sumners, The role of knot theory in DNA research, Boca Raton: CRC Press, 1986. |
| [9] |
T. Schlick, Q. Y. Zhu, A. Dey, S. Jain, S. T. Yan, A. Laederach, To knot or not to knot: multiple conformations of the SARS-CoV-2 frameshifting RNA element, J. Am. Chem. Soc., 143 (2021), 11404–11422. https://dx.doi.org/10.1021/jacs.1c03003 doi: 10.1021/jacs.1c03003
|
| [10] |
K. C. Millett, E. J. Rawdon, A. Stasiak, J. I. Sułkowska, Identifying knots in proteins, Biochem. Soc. Trans., 41 (2013), 533–537. https://dx.doi.org/10.1042/bst20120339 doi: 10.1042/bst20120339
|
| [11] |
J. Qin, S. T. Milner, Counting polymer knots to find the entanglement length, Soft Matter, 7 (2011), 10676–10693. https://dx.doi.org/10.1039/c1sm05972f doi: 10.1039/c1sm05972f
|
| [12] |
Y. Z. Liu, M. O'Keeffe, M. M. J. Treacy, O. M. Yaghi, The geometry of periodic knots, polycatenanes and weaving from a chemical perspective: a library for reticular chemistry, Chem. Soc. Rev., 47 (2018), 4642–4664. https://dx.doi.org/10.1039/c7cs00695k doi: 10.1039/c7cs00695k
|
| [13] |
R. L. Ricca, Topology bounds energy of knots and links, Proc. R. Soc. A., 464 (2008), 293–300. https://dx.doi.org/10.1098/rspa.2007.0174 doi: 10.1098/rspa.2007.0174
|
| [14] |
E. Panagiotou, K. C. Millett, P. J. Atzberger, Topological methods for polymeric materials: characterizing the relationship between polymer entanglement and viscoelasticity, Polymers, 11 (2019), 437. https://dx.doi.org/10.3390/polym11030437 doi: 10.3390/polym11030437
|
| [15] |
J. Arsuaga, M. Vazquez, P. McGuirk, S. Trigueros, D. W. Sumners, J. Roca, DNA knots reveal a chiral organization of DNA in phage capsids, Proc. Natl. Acad. Sci. U.S.A., 102 (2005), 9165–9169. https://dx.doi.org/10.1073/pnas.0409323102 doi: 10.1073/pnas.0409323102
|
| [16] |
J. I. Sulkowska, E. J. Rawdon, K. C. Millet, J. N. Onuchic, A. Stasiak, Conservation of complex knotting and slipknotting patterns in proteins, Proc. Natl. Acad. Sci. U.S.A., 109 (2012), E1715–E1723. https://dx.doi.org/10.1073/pnas.1205918109 doi: 10.1073/pnas.1205918109
|
| [17] |
P. Freyd, D. Yetter, J. Hoste, W. B. R. Lickorish, K. Millett, A. Ocneanu, A new polynomial invariant of knots and links, Bull. Amer. Math. Soc., 12 (1985), 239–246. https://dx.doi.org/10.1090/s0273-0979-1985-15361-3 doi: 10.1090/s0273-0979-1985-15361-3
|
| [18] |
L. H. Kauffman, An invariant of regular isotopy, Trans. Amer. Math. Soc., 318 (1990), 417–471. https://dx.doi.org/10.1090/S0002-9947-1990-0958895-7 doi: 10.1090/S0002-9947-1990-0958895-7
|
| [19] |
J. H. Przytycki, P. Traczyk, Conway algebras and skein equivalence of links, Proc. Amer. Math. Soc., 100 (1987), 744–748. https://dx.doi.org/10.1090/S0002-9939-1987-0894448-2 doi: 10.1090/S0002-9939-1987-0894448-2
|
| [20] |
L. Shen, H. S. Feng, F. L. Li, F. C. Lei, J. Wu, G.-W. Wei, Knot data analysis using multiscale Gauss link integral, Proc. Natl. Acad. Sci. U.S.A., 121 (2024), e2408431121. https://dx.doi.org/10.1073/pnas.2408431121 doi: 10.1073/pnas.2408431121
|
| [21] | E. Panagiotou, K. W. Plaxco, A topological study of protein folding kinetics, 2018, arXiv: 1812.08721. |
| [22] |
Q. Baldwin, E. Panagiotou, The local topological free energy of proteins, J. Theor. Biol., 529 (2021), 110854. https://dx.doi.org/10.1016/j.jtbi.2021.110854 doi: 10.1016/j.jtbi.2021.110854
|
| [23] |
Q. Baldwin, B. Sumpter, E. Panagiotou, The local topological free energy of the SARS-CoV-2 Spike protein, Polymers, 14 (2022), 3014. https://dx.doi.org/10.3390/polym14153014 doi: 10.3390/polym14153014
|
| [24] |
E. Panagiotou, L. H. Kauffman, Knot polynomials of open and closed curves, Proc. R. Soc. A., 476 (2020), 20200124. https://dx.doi.org/10.1098/rspa.2020.0124 doi: 10.1098/rspa.2020.0124
|
| [25] |
K. Barkataki, E. Panagiotou, The Jones polynomial of collections of open curves in 3-space, Proc. R. Soc. A., 478 (2022), 20220302. https://dx.doi.org/10.1098/rspa.2022.0302 doi: 10.1098/rspa.2022.0302
|
| [26] |
E. Panagiotou, L. H. Kauffman, Vassiliev measures of complexity of open and closed curves in 3-space, Proc. R. Soc. A., 477 (2021), 20210440. https://dx.doi.org/10.1098/rspa.2021.0440 doi: 10.1098/rspa.2021.0440
|
| [27] |
J. Wang, E. Panagiotou, The protein folding rate and the geometry and topology of the native state, Sci. Rep., 12 (2022), 6384. https://dx.doi.org/10.1038/s41598-022-09924-0 doi: 10.1038/s41598-022-09924-0
|
| [28] |
T. Herschberg, K. Pifer, E. Panagiotou, A computational package for measuring Topological Entanglement in Polymers, Proteins and Periodic systems (TEPPP), Comput. Phys. Commun., 286 (2023), 108639. https://dx.doi.org/10.1016/j.cpc.2022.108639 doi: 10.1016/j.cpc.2022.108639
|
| [29] |
Z. X. Cang, G.-W. Wei, Persistent cohomology for data with multicomponent heterogeneous information, SIAM J. Math. Data Sci., 2 (2020), 396–418. https://dx.doi.org/10.1137/19m1272226 doi: 10.1137/19m1272226
|
| [30] |
J. K. Park, R. Jernigan, Z. J. Wu, Coarse grained normal mode analysis vs. refined gaussian network model for protein residue-level structural fluctuations, Bull. Math. Biol., 75 (2013), 124–160. https://dx.doi.org/10.1007/s11538-012-9797-y doi: 10.1007/s11538-012-9797-y
|
| [31] | V. Turaev, Knotoids, Osaka J. Math., 49 (2012), 195–223. https://dx.doi.org/10.18910/10080 |
| [32] | N. Gügümcü, L. H. Kauffman, New invariants of knotoids, Eur. J. Combin., 65 (2017), 186–229. https://dx.doi.org/10.1016/j.ejc.2017.06.004 |
| [33] | N. Gügümcü, S. Lambropoulou, Knotoids, braidoids and applications, Symmetry, 9 (2017), 315. https://dx.doi.org/10.3390/sym9120315 |
| [34] | N. Gügümcü, L. Kauffman, Parity in knotoids, 2019, arXiv: 1905.04089. |
| [35] |
M. Manouras, S. Lambropoulou, L. H. Kauffman, Finite type invariants for knotoids, Eur. J. Combin., 98 (2021), 103402. https://dx.doi.org/10.1016/j.ejc.2021.103402 doi: 10.1016/j.ejc.2021.103402
|
| [36] | D. Cohen-Steiner, H. Edelsbrunner, J. Harer, Stability of persistence diagrams, In: Proceedings of the twenty-first annual symposium on computational geometry, New York: Association for Computing Machinery, 2005,263–271. https://dx.doi.org/10.1145/1064092.1064133 |
| [37] |
Z. X. Cang, E. Munch, G.-W. Wei, Evolutionary homology on coupled dynamical systems with applications to protein flexibility analysis, J. Appl. and Comput. Topology, 4 (2020), 481–507. https://dx.doi.org/10.1007/s41468-020-00057-9 doi: 10.1007/s41468-020-00057-9
|
| [38] |
D. Bramer, G.-W. Wei, Atom-specific persistent homology and its application to protein flexibility analysis, Comput. Math. Biophys., 8 (2020), 1–35. https://dx.doi.org/10.1515/cmb-2020-0001 doi: 10.1515/cmb-2020-0001
|
| [39] |
K. Opron, K. L. Xia, G.-W. Wei, Fast and anisotropic flexibility-rigidity index for protein flexibility and fluctuation analysis, J. Chem. Phys., 140 (2014), 234105. https://dx.doi.org/10.1063/1.4882258 doi: 10.1063/1.4882258
|







Figures(8)

Ruzhi Song, Fengling Li, Jie Wu, Fengchun Lei, Guo-Wei Wei. Multi-scale Jones polynomial and persistent Jones polynomial for knot data analysis[J]. AIMS Mathematics, 2025, 10(1): 1463-1487. doi: 10.3934/math.2025068







 DownLoad:
DownLoad: