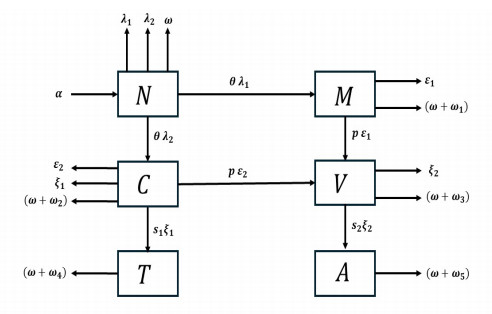
The relationship between cancer and the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection is controversial. While SARS-CoV-2 can worsen the status of a cancer patient, many remission cases after SARS-CoV-2 infection have been recorded. It has been suggested that SARS-CoV-2 could have oncolytic properties, which needs further investigations. Mathematical modeling is a powerful tool that can significantly enhance experimental and medical studies. Our objective was to propose and analyze a mathematical model for oncolytic SARS-CoV-2 with immunity. The basic properties of this model, including existence, uniqueness, nonnegativity, and boundedness of the solutions, were confirmed. The equilibrium points were computed, and their existence conditions were determined. The global stability of the equilibria was proven using the Lyapunov theory. Numerical simulations were implemented to validate the theoretical results. It was found that the model has thirteen equilibrium points that reflect different infection states. Based on the model's results, the infection of cancer cells by SARS-CoV-2 can lead to a reduction in the concentration of cancer cells. Additionally, the induction of cytotoxic T lymphocytes (CTLs) decreases the number of cancer cells, potentially resulting in cancer remission or an improvement in the overall health of cancer patients. This theoretical result aligns with numerous studies highlighting the oncolytic role of SARS-CoV-2. In addition, given the limited availability of real data, further studies are essential to better comprehend the role of immune responses and their impact on the oncolytic role of SARS-CoV-2.
Citation: Afnan Al Agha, Hakim Al Garalleh. Oncolysis by SARS-CoV-2: modeling and analysis[J]. AIMS Mathematics, 2024, 9(3): 7212-7252. doi: 10.3934/math.2024351
The relationship between cancer and the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection is controversial. While SARS-CoV-2 can worsen the status of a cancer patient, many remission cases after SARS-CoV-2 infection have been recorded. It has been suggested that SARS-CoV-2 could have oncolytic properties, which needs further investigations. Mathematical modeling is a powerful tool that can significantly enhance experimental and medical studies. Our objective was to propose and analyze a mathematical model for oncolytic SARS-CoV-2 with immunity. The basic properties of this model, including existence, uniqueness, nonnegativity, and boundedness of the solutions, were confirmed. The equilibrium points were computed, and their existence conditions were determined. The global stability of the equilibria was proven using the Lyapunov theory. Numerical simulations were implemented to validate the theoretical results. It was found that the model has thirteen equilibrium points that reflect different infection states. Based on the model's results, the infection of cancer cells by SARS-CoV-2 can lead to a reduction in the concentration of cancer cells. Additionally, the induction of cytotoxic T lymphocytes (CTLs) decreases the number of cancer cells, potentially resulting in cancer remission or an improvement in the overall health of cancer patients. This theoretical result aligns with numerous studies highlighting the oncolytic role of SARS-CoV-2. In addition, given the limited availability of real data, further studies are essential to better comprehend the role of immune responses and their impact on the oncolytic role of SARS-CoV-2.

| [1] | World Health Organization, COVID-19 epidemiological update - 27 October 2023, 2023. Available from: https://www.who.int/publications/m/item/covid-19-epidemiological-update---27-october-2023. |
| [2] |
S. A. Gold, V. Margulis, Uncovering a link between COVID-19 and renal cell carcinoma, Nat. Rev. Urol., 20 (2023), 330–331. https://doi.org/10.1038/s41585-023-00749-8 doi: 10.1038/s41585-023-00749-8

|
| [3] |
M. Shariq, J. A. Sheikh, N. Quadir, N. Sharma, S. E. Hasnain, N. Z. Ehtesham, COVID-19 and tuberculosis: the double whammy of respiratory pathogens, Eur. Respir. Rev., 31 (2022), 210264. https://doi.org/10.1183/16000617.0264-2021 doi: 10.1183/16000617.0264-2021
|
| [4] |
A. Tahmasebzadeh, R. Paydar, H. Kaeidi, Lifetime attributable breast cancer risk related to lung CT scan in women with Covid19, Front. Biomed. Technol., 11 (2024), 69–74. https://doi.org/10.18502/fbt.v11i1.14513 doi: 10.18502/fbt.v11i1.14513
|
| [5] |
Y. S. Li, H. C. Ren, J. H. Cao, Correlation of SARS‑COV‑2 to cancer: carcinogenic or anticancer? (review), Int. J. Oncolytics, 60 (2022), 42. https://doi.org/10.3892/ijo.2022.5332 doi: 10.3892/ijo.2022.5332
|
| [6] |
C. Meo, G. Palma, F. Bruzzese, A. Budillon, C. Napoli, F. de Nigris, Spontaneous cancer remission after COVID-19: insights from the pandemic and their relevance for cancer treatment, J. Transl. Med., 21 (2023), 273. https://doi.org/10.1186/s12967-023-04110-w doi: 10.1186/s12967-023-04110-w
|
| [7] |
M. Costanzo, M. A. R. de Giglio, G. N. Roviello, Deciphering the relationship between SARS-COV-2 and cancer, Int. J. Mol. Sci., 24 (2023), 7803. https://doi.org/10.3390/ijms24097803 doi: 10.3390/ijms24097803
|
| [8] |
H. Goubran, J. Stakiw, J. Seghatchian, G. Ragab, T. Burnouf, SARS-COV-2 and cancer: the intriguing and informative cross-talk, Transfus. Apher. Sci., 61 (2022), 103488. https://doi.org/10.1016/j.transci.2022.103488 doi: 10.1016/j.transci.2022.103488
|
| [9] |
D. Barh, S. Tiwari, L. G. R. Gomes, M. E. Weener, K. J. Alzahrani, K. Alsharif, et al., Potential molecular mechanisms of rare anti-tumor immune response by SARS-COV-2 in isolated cases of lymphomas, Viruses, 13 (2021), 1927. https://doi.org/10.3390/v13101927 doi: 10.3390/v13101927
|
| [10] |
O. K. Choong, R. Jakobsson, A. G. Bergdahl, S. Brunet, A. Kärmander, J. Waldenström, et al., SARS-COV-2 replicates and displays oncolytic properties in clear cell and papillary renal cell carcinoma, PLoS One, 18 (2023), e0279578. https://doi.org/10.1371/journal.pone.0279578 doi: 10.1371/journal.pone.0279578
|
| [11] |
E. J. Schafer, F. Islami, X. Han, L. M. Nogueira, N. S. Wagle, K. R. Yabroff, et al., Changes in cancer incidence rates by stage during the COVID-19 pandemic in the US, Int. J. Cancer, 154 (2024), 786–792. https://doi.org/10.1002/ijc.34758 doi: 10.1002/ijc.34758
|
| [12] |
J. P. Bounassar-Filho, L. Boeckler-Troncoso, J. Cajigas-Gonzalez, M. G. Zavala-Cerna, SARS-COV-2 as an oncolytic virus following reactivation of the immune system: a review, Int. J. Mol. Sci., 24 (2023), 2326. https://doi.org/10.3390/ijms24032326 doi: 10.3390/ijms24032326
|
| [13] |
D. H. Shin, A. Gillard, A. V. Wieren, C. Gomez-Manzano, J. Fueyo, Remission of liquid tumors and SARS-COV-2 infection: a literature review, Mol. Ther. Oncolytics, 26 (2022), 135–140. https://doi.org/10.1016/j.omto.2022.06.006 doi: 10.1016/j.omto.2022.06.006
|
| [14] | A. Donia, R. Shahid, M. Nawaz, T. Yaqub, H. Bokhari, Can we develop oncolytic SARSCOV-2 to specifically target cancer cells? Ther. Adv. Med. Oncolytics, 13 (2021), 1988. https://doi.org/10.1177/17588359211061988 |
| [15] |
F. Pasin, M. M. Calveri, A. Calabrese, G. Pizzarelli, I. Bongiovanni, M. Andreoli, et al., Oncolytic effect of SARS-CoV-2 in a patient with NK lymphoma, Acta Biomed., 91 (2020), 10141. https://doi.org/10.23750/abm.v91i3.10141 doi: 10.23750/abm.v91i3.10141
|
| [16] |
S. Challenor, D. Tucker, SARS‐COV‐2‐induced remission of Hodgkin lymphoma, Br. J. Haematol., 192 (2021), 415. https://doi.org/10.1111/bjh.17116 doi: 10.1111/bjh.17116
|
| [17] |
M. Sollini, F. Gelardi, C. Carlo-Stella, A. Chiti, Complete remission of follicular lymphoma after SARS-COV-2 infection: from the "flare phenomenon" to the "abscopal effect", Eur. J. Nucl. Med. Mol. Imaging, 48 (2021), 2652–2654. https://doi.org/10.1007/s00259-021-05275-6 doi: 10.1007/s00259-021-05275-6
|
| [18] | E. Z. Kandeel, L. Refaat, R. Abdel-Fatah, M. Samra, A. Bayoumi, M. S. Abdellateif, et al., Could COVID-19 induce remission of acute leukemia? Hematology, 26 (2021), 870–873. https://doi.org/10.1080/16078454.2021.1992117 |
| [19] |
D. Antwi-Amoabeng, M. B. Ulanja, B. D. Beutler, S. V. Reddy, Multiple myeloma remission following COVID-19: an observation in search of a mechanism (a case report), Pan Afr. Med. J., 39 (2021), 117. https://doi.org/10.11604/pamj.2021.39.117.30000 doi: 10.11604/pamj.2021.39.117.30000
|
| [20] |
L. Ohadi, F. Hosseinzadeh, S. Dadkhahfar, S. Nasiri, Oncolytic effect of SARS-CoV-2 in a patient with mycosis fungoides: a case report, Clin. Case Rep., 10 (2022), e05682. https://doi.org/10.1002/ccr3.5682 doi: 10.1002/ccr3.5682
|
| [21] | P. Agarwal, J. Nieto, D. Torres, Mathematical analysis of infectious diseases, Academic Press, 2022. |
| [22] |
A. Debbouche, J. J. Nieto, D. F. M. Torres, Focus point: cancer and HIV/AIDS dynamics-from optimality to modellin, Eur. Phys. J. Plus, 136 (2021), 165. https://doi.org/10.1140/epjp/s13360-021-01154-z doi: 10.1140/epjp/s13360-021-01154-z
|
| [23] |
O. Nave, I. Hartuv, U. Shemesh, $\Theta$-SEIHRD mathematical model of Covid19-stability analysis using fast-slow decomposition, PeerJ, 8 (2020), e10019. https://doi.org/10.7717/peerj.10019 doi: 10.7717/peerj.10019
|
| [24] |
A. Atifa, M. A. Khan, K. Iskakova, F. S. Al-Duais, I. Ahmad, Mathematical modeling and analysis of the SARS-Cov-2 disease with reinfection, Comput. Biol. Chem., 98 (2022), 107678. https://doi.org/10.1016/j.compbiolchem.2022.107678 doi: 10.1016/j.compbiolchem.2022.107678
|
| [25] | J. H. Rojas, M. Paredes, M. Banerjee, O. Akman, A. Mubayi, Mathematical modeling and dynamics of SARS-COV-2 in Colombia, Lett. Biomath., 9 (2022), 41-–56. |
| [26] |
O. Nave, M. Sigron, A mathematical model for cancer treatment based on combination of anti-angiogenic and immune cell therapies, Results Appl. Math., 16 (2022), 10030. https://doi.org/10.1016/j.rinam.2022.100330 doi: 10.1016/j.rinam.2022.100330
|
| [27] |
A. M. Elaiw, A. D. A. Agha, Global dynamics of SARS-CoV-2/cancer model with immune responses, Appl. Math. Comput., 408 (2021), 126364. https://doi.org/10.1016/j.amc.2021.126364 doi: 10.1016/j.amc.2021.126364
|
| [28] |
A. M. Elaiw, A. D. Hobiny, A. D. A. Agha, Global dynamics of reaction-diffusion oncolytic M1 virotherapy with immune response, Appl. Math. Comput., 367 (2020), 124758. https://doi.org/10.1016/j.amc.2019.124758 doi: 10.1016/j.amc.2019.124758
|
| [29] |
J. Malinzi, P. Sibanda, H. Mambili-Mamboundou, Analysis of virotherapy in solid tumor invasion, Math. Biosci., 263 (2015), 102–110. https://doi.org/10.1016/j.mbs.2015.01.015 doi: 10.1016/j.mbs.2015.01.015
|
| [30] |
T. Alzahrani, R. Eftimie, D. Trucu, Multiscale modelling of cancer response to oncolytic viral therapy, Math. Biosci., 310 (2019), 76–95. https://doi.org/10.1016/j.mbs.2018.12.018 doi: 10.1016/j.mbs.2018.12.018
|
| [31] | K. W. Okamoto, P. Amarasekare, I. T. D. Petty, Modeling oncolytic virotherapy: is complete tumor-tropism too much of a good thing? J. Theor. Biol., 358 (2014), 166–178. https://doi.org/10.1016/j.jtbi.2014.04.030 |
| [32] |
J. Zhao, J. P. Tian, Spatial model for oncolytic virotherapy with lytic cycle delay, Bull. Math. Biol., 81 (2019), 2396–2427. https://doi.org/10.1007/s11538-019-00611-2 doi: 10.1007/s11538-019-00611-2
|
| [33] |
Z. Wang, Z. Guo, H. Peng, A mathematical model verifying potent oncolytic efficacy of M1 virus, Math. Biosci., 276 (2016), 19–27. https://doi.org/10.1016/j.mbs.2016.03.001 doi: 10.1016/j.mbs.2016.03.001
|
| [34] | R. G. Bartle, D. R. Sherbert, Introduction to real analysis, John Wiley & Sons, Inc., 2000. |
| [35] | E. Jones, P. Romemer, M. Raghupathi, S. Pankavich, Analysis and simulation of the three-component model of HIV dynamics, arXiv, 2013. https://doi.org/10.48550/arXiv.1312.3671 |
| [36] | H. L. Smith, P. Waltman, The theory of the chemostat: dynamics of microbial competition, Cambridge University Press, 1995. https://doi.org/10.1017/CBO9780511530043 |
| [37] | H. K. Khalil, Nonlinear Systems, Prentice-Hall, 1996. |
| [38] |
T. Sumi, K. Harada, Immune response to SARS-CoV-2 in severe disease and long COVID-19, iScience, 25 (2022), 104723. https://doi.org/10.1016/j.isci.2022.104723 doi: 10.1016/j.isci.2022.104723
|
| [39] |
C. Li, J. Xu, J. Liu, Y. Zhou, The within-host viral kinetics of SARS-CoV-2, Math. Biosci. Eng., 17 (2020), 2853–2861. https://doi.org/10.3934/mbe.2020159 doi: 10.3934/mbe.2020159
|










Figures(11) / Tables(1)

Afnan Al Agha, Hakim Al Garalleh. Oncolysis by SARS-CoV-2: modeling and analysis[J]. AIMS Mathematics, 2024, 9(3): 7212-7252. doi: 10.3934/math.2024351







 DownLoad:
DownLoad: