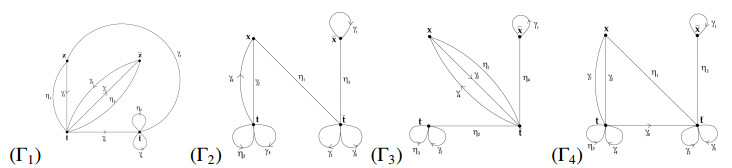
It was demonstrated by Bibi and Edjvet in [
Citation: Mairaj Bibi, Sajid Ali, Muhammad Shoaib Arif, Kamaleldin Abodayeh. Solving singular equations of length eight over torsion-free groups[J]. AIMS Mathematics, 2023, 8(3): 6407-6431. doi: 10.3934/math.2023324
It was demonstrated by Bibi and Edjvet in [

| [1] |
M. Bibi, M. Edjvet, Solving equations of length seven over torsion-free groups, J. Group Theory, 21 (2018), 147–164. https://doi.org/10.1515/jgth-2017-0032 doi: 10.1515/jgth-2017-0032

|
| [2] |
F. Levin, Solutions of equations over groups, Bull. Amer. Math. Soc., 68 (1962), 603–604. https://doi.org/10.1090/S0002-9904-1962-10868-4 doi: 10.1090/S0002-9904-1962-10868-4
|
| [3] |
M. I. Prischschipov, On small length equations over torsion-free groups, Int. J. Algebr. Comput., 4 (1994), 575–589. https://doi.org/10.1142/S0218196794000166 doi: 10.1142/S0218196794000166
|
| [4] |
W. A. Bogley, S. J. Pride, Aspherical relative presentations, P. Edinburgh Math. Soc., 35 (1992), 1–39. https://doi.org/10.1017/S0013091500005290 doi: 10.1017/S0013091500005290
|
| [5] | R. C. Lyndon, P. E. Schupp, Combinatorial group theory, Springer-Verlag, Heidelberg, 1977. |
| [6] |
S. V. Ivanov, A. A. Klyachko, Solving equations of length at most six over torsion-free groups, J. Group Theory, 3 (2006), 329–337. https://doi.org/10.1515/jgth.2000.026 doi: 10.1515/jgth.2000.026
|
| [7] | J. R. Stallings, A graph-theoretic lemma and group-embeddings, Combinatorial Group Theory and Topology, Princeton University Press, 111 (2016), 145–156. |
| [8] |
J. Howie, On pairs of 2-complexes and systems of equations over groups, J. Reine Angew. Math., 324 (1981), 165–174. https://doi.org/10.1515/crll.1981.324.165 doi: 10.1515/crll.1981.324.165
|
| [9] |
M. Gerstenhaber, D. S. Rothaus, The solution of sets of equations over groups, P. Natl. A. Sci. USA, 48 (1962), 1531–1533. https://doi.org/10.1073/pnas.48.9.1531 doi: 10.1073/pnas.48.9.1531
|
| [10] | A. Evangelidou, The solution of length five equations over groups, Commun. Algebra, 35 (2007). https://doi.org/10.1080/00927870701247039 |
| [11] |
M. Edjvet, On the asphericity of one relator relative presentations, P. Roy. Soc. Edinb., 124 (1994), 713–728. https://doi.org/10.1017/S0308210500028614 doi: 10.1017/S0308210500028614
|
| [12] |
J. Howie, On the asphericity of ribbon disc complements, Trans. Amer. Math. Soc., 289 (1985), 281–302. https://doi.org/10.1090/S0002-9947-1985-0779064-8 doi: 10.1090/S0002-9947-1985-0779064-8
|
| [13] |
A. Clifford, R. Z. Goldstein, Equations with torsion-free coefficients, P. Edinb. Math. Soc., 43 (2000), 295–307. https://doi.org/10.1017/S0013091500020939 doi: 10.1017/S0013091500020939
|
| [14] |
A. A. Klyachko, A funny property of sphere and equations over groups, Commun. Algebra, 21 (1993), 2555–2575. https://doi.org/10.1080/00927879308824692 doi: 10.1080/00927879308824692
|
| [15] |
S. J. Pride, Involutary presentations with applications to Coxeter groups, NEC-groups and groups of Kanevskiĭ, J. Algebra, 120 (1989), 200–223. https://doi.org/10.1016/0021-8693(89)90195-6 doi: 10.1016/0021-8693(89)90195-6
|
| [16] |
J. Huebschmann, Cohomology theory of aspherical groups and of small cancellation groups, J. Pure Appl. Algebra, 14 (1979), 137–143. https://doi.org/10.1016/0022-4049(79)90003-3 doi: 10.1016/0022-4049(79)90003-3
|
| [17] |
M. Edjvet, Equations over groups and a theorem of Higman, Neumann, and Neumann, P. London Math. Soc., 62 (1991), 563–589. https://doi.org/10.1112/plms/s3-62.3.563 doi: 10.1112/plms/s3-62.3.563
|
| [18] | W. A. Bogley, Local collapses for diagrammatic reducibility, Topology and Combinatorial Group Theory, Lecture Notes in Mathematics, Springer, Berlin, Heidelberg, 1440 (1990). https://doi.org/10.1007/BFb0084447 |
| [19] |
S. G. Brick, Normal-convexity and equations over groups, Invent. Math., 94 (1988), 81–104. https://doi.org/10.1007/BF01394345 doi: 10.1007/BF01394345
|
| [20] | R. Brown, J. Huebschmann, Identities among relations in low-dimensional topology, London Mathematical Society Lecture Note Series, 48 (1982). |
| [21] |
D. J. Collins, J. Huebschmann, Spherical diagrams and identities among relations, Math. Ann., 261 (1982), 155–183. https://doi.org/10.1007/BF01456216 doi: 10.1007/BF01456216
|
| [22] |
D. J. Collins, J. Perraud, Cohomology and finite subgroups of small cancellation quotients of free products, Math. Proc. Cambridge Philos. Soc., 97 (1985), 243–259. https://doi.org/10.1017/S0305004100062800 doi: 10.1017/S0305004100062800
|
| [23] |
E. J. Fennessey, S. J. Pride, Equivalences of two-complexes, with applications to NEC-groups, Math. Proc. Cambridge Philos. Soc., 106 (1989), 215–228. https://doi.org/10.1017/S0305004100078038 doi: 10.1017/S0305004100078038
|
| [24] | S. M. Gersten, Reducible diagrams and equations over groups, Essays in Group Theory, Mathematical Sciences Research Institute Publications, Springer, New York, 8 (1987). https://doi.org/10.1007/978-1-4613-9586-7_2 |














Figures(15)

Mairaj Bibi, Sajid Ali, Muhammad Shoaib Arif, Kamaleldin Abodayeh. Solving singular equations of length eight over torsion-free groups[J]. AIMS Mathematics, 2023, 8(3): 6407-6431. doi: 10.3934/math.2023324







 DownLoad:
DownLoad: