A novel scheme based on the generalized differential operator and computer algebra was used to construct solitary solutions to a system of Riccati differential equations with diffusive coupling. The presented approach yields necessary and sufficient existence conditions of solitary solutions with respect to the system parameters. The proposed stepwise approach enabled the derivation of the explicit analytic solution, which could not be derived using direct balancing techniques due to the complexity of algebraic relationships. Computational experiments were used to demonstrate the efficacy of proposed scheme.
Citation: Romas Marcinkevicius, Inga Telksniene, Tadas Telksnys, Zenonas Navickas, Minvydas Ragulskis. The step-wise construction of solitary solutions to Riccati equations with diffusive coupling[J]. AIMS Mathematics, 2023, 8(12): 30683-30703. doi: 10.3934/math.20231568
A novel scheme based on the generalized differential operator and computer algebra was used to construct solitary solutions to a system of Riccati differential equations with diffusive coupling. The presented approach yields necessary and sufficient existence conditions of solitary solutions with respect to the system parameters. The proposed stepwise approach enabled the derivation of the explicit analytic solution, which could not be derived using direct balancing techniques due to the complexity of algebraic relationships. Computational experiments were used to demonstrate the efficacy of proposed scheme.

| [1] |
N. J. Zabusky, M. D. Kruskal, Interaction of "solitons" in a collisionless plasma and the recurrence of initial states, Phys. Rev. Lett., 15 (1965), 240. https://doi.org/10.1103/PhysRevLett.15.240 doi: 10.1103/PhysRevLett.15.240

|
| [2] | L. Munteanu, S. Donescu, Introduction to soliton theory: Applications to mechanics, vol. 143, Springer Science & Business Media, 2004. |
| [3] | A. S. Davydov, Solitons in molecular systems, Springer, 1985. |
| [4] | N. Akhmediev, A. Ankiewicz, Dissipative solitons: from optics to biology and medicine, vol. 751, Springer Science & Business Media, 2008. |
| [5] |
A. S. Johnson, W. Winlow, The soliton and the action potential–primary elements underlying sentience, Front. Physiol., 9 (2018), 779. https://doi.org/10.1039/C8SC90018C doi: 10.1039/C8SC90018C
|
| [6] | A. R. Seadawy, H. Zahed, M. Iqbal, Solitary wave solutions for the higher dimensional Jimo-Miwa dynamical equation via new mathematical techniques, Mathematics, 10 (2022), 1011. |
| [7] | B. Halder, S. Ghosh, P. Basu, J. Bera, B. Malomed, U. Roy, Exact solutions for solitary waves in a Bose-Einstein condensate under the action of a four-color optical lattice, Symmetry, 14 (2021), 49. |
| [8] | X. Liu, The stability of exact solitary wave solutions for simplified modified Camassa–Holm equation, Communications in Nonlinear Science and Numerical Simulation, 106224. |
| [9] |
G. Slavcheva, M. V. Koleva, A. Pimenov, The impact of microcavity wire width on polariton soliton existence and multistability, J. Optics, 19 (2017), 065404. https://doi.org/10.1088/2040-8986/aa6d40 doi: 10.1088/2040-8986/aa6d40
|
| [10] |
T. Han, Z. Li, K. Shi, G. C. Wu, Bifurcation and traveling wave solutions of stochastic Sanakov model with multiplicative white noise in birefringent fibers, Chaos, Soliton. Fract., 163 (2022), 112548. https://doi.org/10.1016/j.chaos.2022.112548 doi: 10.1016/j.chaos.2022.112548
|
| [11] |
T. Han, Z. Li, X. Zhang, Bifurcation and new exact traveling wave solutions to time-space coupled fractional nonlinear Schrödinger equation, Phys. Lett. A, 395 (2021), 127217. https://doi.org/10.1016/j.physleta.2021.127217 doi: 10.1016/j.physleta.2021.127217
|
| [12] |
T. Han, Z. Li. C. Li, Bifurcation analysis, stationary optical solitons and exact solutions for generalized nonlinear Schrödinger equation with nonlinear chromatic dispersion and quintuple power-law of refractive index in optical fibers, Physica A, 615 (2023), 128599. https://doi.org/10.1016/j.physa.2023.128599 doi: 10.1016/j.physa.2023.128599
|
| [13] |
T. Han, Z. Li, J. Yuan, Optical solitons and single traveling wave solutions of Biswas-Arshed equation in birefringent fibers with the beta-time derivative, AIMS Math., 7 (2022), 15282–15297. https://doi.org/10.3934/math.2022837 doi: 10.3934/math.2022837
|
| [14] |
T. Han, Z. Li, K. Zhang, Exact solutions of the stochastic fractional long–short wave interaction system with multiplicative noise in generalized elastic medium, Results Phys., 44 (2023), 106174. https://doi.org/10.1016/j.rinp.2022.106174 doi: 10.1016/j.rinp.2022.106174
|
| [15] | S. Cui, Z. Wang, J. Han, X. Cui, Q. Meng, A deep learning method for solving high-order nonlinear soliton equations, Communications in Theoretical Physics. |
| [16] |
R. Zheng, Z. Yin, Wave breaking and solitary wave solutions for a generalized Novikov equation, Appl. Math. Lett., 100 (2020), 106014. https://doi.org/10.1016/j.rinp.2022.106174 doi: 10.1016/j.rinp.2022.106174
|
| [17] |
O. Nikan, Z. Avazzadeh, M. Rasoulizadeh, Soliton solutions of the nonlinear sine-Gordon model with Neumann boundary conditions arising in crystal dislocation theory, Nonlinear Dynam., 106 (2021), 783–813. https://doi.org/10.1007/s11071-021-06822-4 doi: 10.1007/s11071-021-06822-4
|
| [18] | M. Sciacca, I. Carlomagno, A. Sellitto, Thermal solitons in nanotubes, Wave Motion, 102967. |
| [19] | S. H. Dong, Schrödinger equation with the potential V(r) = ar-4+ br-3+ cr-2+ dr-1, Phys. Scripta, 64 (2001), 273. |
| [20] |
S. H. Dong, The ansatz method for analyzing Schrödinger's equation with three anharmonic potentials in d dimensions, J. Genet. Couns., 15 (2002), 385–395. https://doi.org/10.1023/A:1021220712636 doi: 10.1023/A:1021220712636
|
| [21] |
M. S. Child, S. H. Dong, X. G. Wang, Quantum states of a sextic potential: hidden symmetry and quantum monodromy, J. Phys. A-Math. Gen., 33 (2000), 5653. https://doi.org/10.1023/A:1021220712636 doi: 10.1023/A:1021220712636
|
| [22] |
Y. S. Guo, W. Li, S. H. Dong, Gaussian solitary solution for a class of logarithmic nonlinear Schrödinger equation in (1+ n) dimensions, Results Phys., 44 (2023), 106187. https://doi.org/10.1016/j.rinp.2022.106187 doi: 10.1016/j.rinp.2022.106187
|
| [23] |
R. C. López, G. H. Sun, O. Camacho-Nieto, C. Yáñez-Márquez, S. H. Dong, Analytical traveling-wave solutions to a generalized Gross–Pitaevskii equation with some new time and space varying nonlinearity coefficients and external fields, Phys. Lett. A, 381 (2017), 2978–2985. https://doi.org/10.1016/j.rinp.2022.106187 doi: 10.1016/j.rinp.2022.106187
|
| [24] |
Z. Navickas, L. Bikulciene, M. Rahula, M. Ragulskis, Algebraic operator method for the construction of solitary solutions to nonlinear differential equations, Commun. Nonlinear Sci. Numer. Simul., 18 (2013), 1374–1389. https://doi.org/10.1016/j.cnsns.2012.10.009 doi: 10.1016/j.cnsns.2012.10.009
|
| [25] |
Z. Navickas, R. Marcinkevicius, T. Telksnys, M. Ragulskis, Existence of second order solitary solutions to Riccati differential equations coupled with a multiplicative term, IMA J. Appl. Math., 81 (2016), 1163–1190. https://doi.org/10.1093/imamat/hxw050 doi: 10.1093/imamat/hxw050
|
| [26] | A. Scott, Eds, Encyclopedia of Nonlinear Science, Routledge, New York, 2004. |
| [27] |
Z. Navickas, T. Telksnys, I. Timofejeva, M. Ragulskis, R. Marcinkevicius, An analytical scheme for the analysis of multi-hump solitons, Adv. Complex Syst., 22 (2019), 1850027. https://doi.org/10.1142/S0219525918500273 doi: 10.1142/S0219525918500273
|
| [28] |
V. L. Kurakin, A. S. Kuzmin, A. V. Mikhalev, A. A. Nechaev, Linear recurring sequences over rings and modules, J. Math. Sci., 76 (1995), 2793–2915. https://doi.org/10.1007/BF02362772 doi: 10.1007/BF02362772
|
| [29] |
D. E. Knuth, Two notes on notation, Am. Math. Mon., 99 (1992), 403–422. https://doi.org/10.1080/00029890.1992.11995869 doi: 10.1080/00029890.1992.11995869
|
| [30] |
N. A. Kudryashov, Seven common errors in finding exact solutions of nonlinear differential equations, Commun. Nonlinear Sci. Numer. Simul., 14 (2009), 3507–3529. https://doi.org/10.1016/j.cnsns.2009.01.023 doi: 10.1016/j.cnsns.2009.01.023
|
| [31] |
R. O. Popovych, O. O. Vaneeva, More common errors in finding exact solutions of nonlinear differential equations: Part Ⅰ, Commun. Nonlinear Sci. Numer. Simul., 15 (2010), 3887–3899. https://doi.org/10.1016/j.cnsns.2010.01.037 doi: 10.1016/j.cnsns.2010.01.037
|
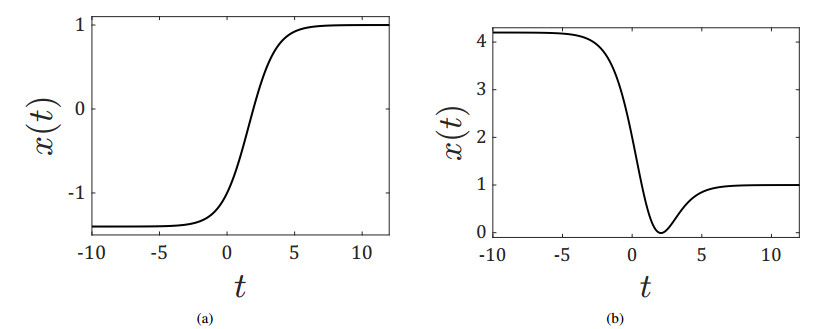
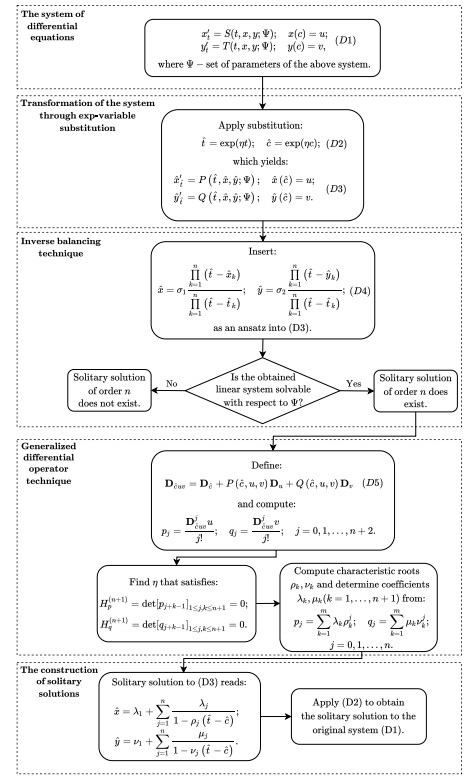
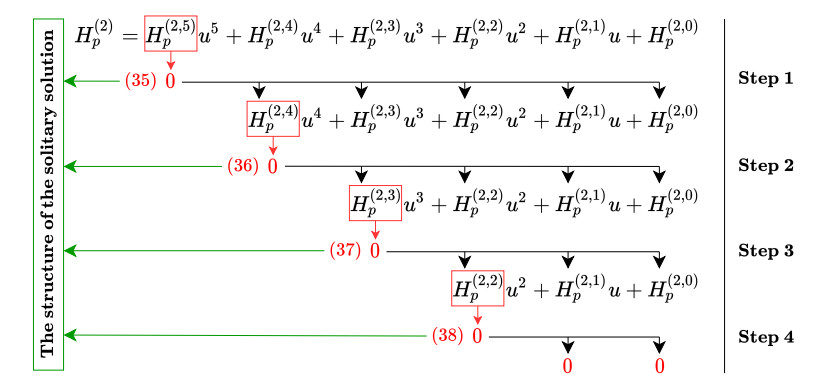
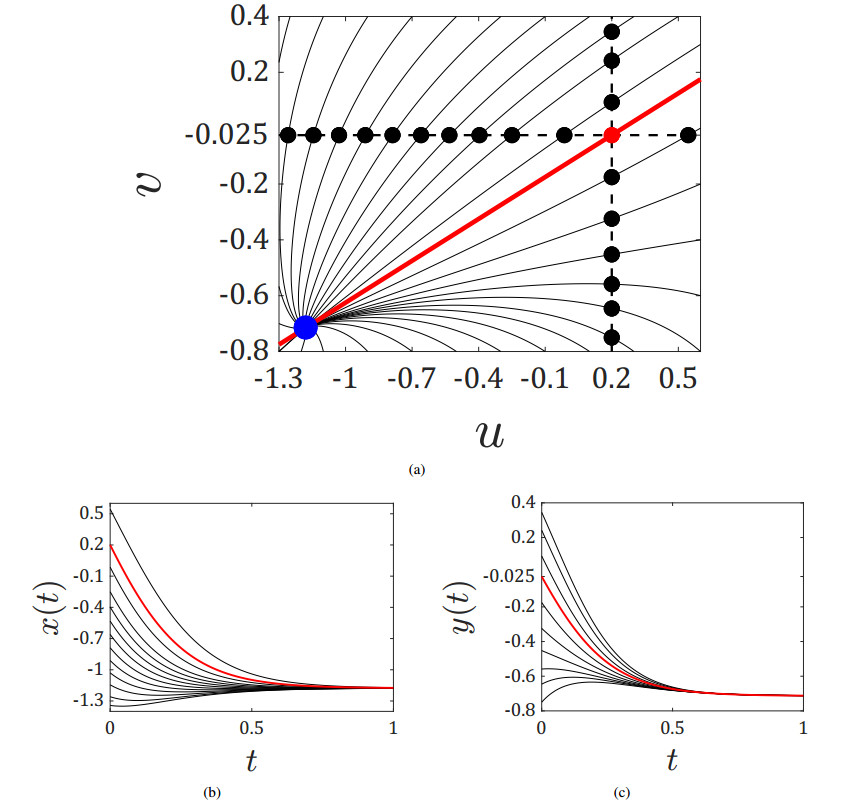
Figures(5)

Romas Marcinkevicius, Inga Telksniene, Tadas Telksnys, Zenonas Navickas, Minvydas Ragulskis. The step-wise construction of solitary solutions to Riccati equations with diffusive coupling[J]. AIMS Mathematics, 2023, 8(12): 30683-30703. doi: 10.3934/math.20231568







 DownLoad:
DownLoad: