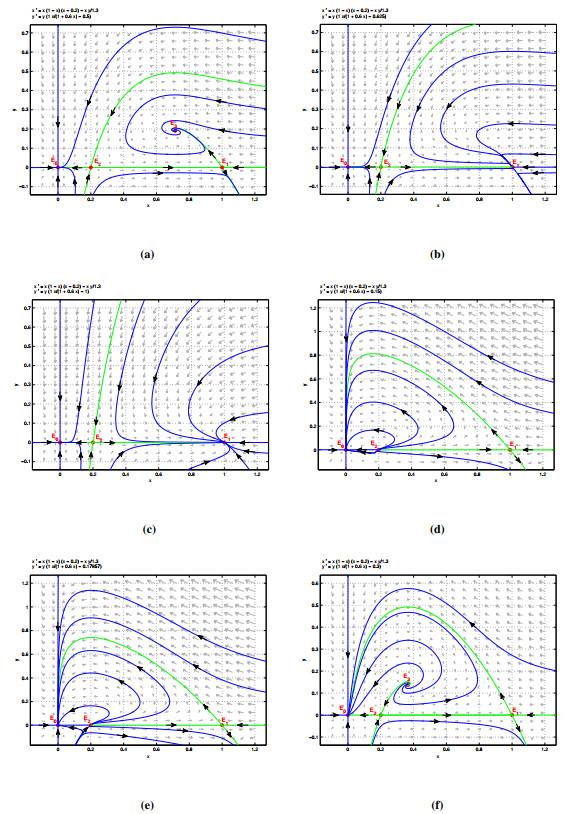
In this work, we propose and investigate a new predator-prey model with strong Allee effect in prey and Holling type Ⅱ functional response in predator. By performing a comprehensive dynamical analysis, we first derive the existence and stability of all the possible equilibria of the system and the system undergoes two transcritical bifurcations and one Hopf-bifurcation. Next, we have calculated the first Lyapunov coefficient and find the Hopf-bifurcation in this model is supercritical and a stable limit cycle is born. Then, by comparing the properties of the system with and without Allee effect, we show that the strong Allee effect is of great importance to the dynamics. It can drive the system to instability. Specifically, Allee effect can increase the extinction risk of populations and has the ability to switch the system's stability to limit cycle oscillation from stable node. Moreover, numerical simulations are presented to prove the validity of our findings.
Citation: Yudan Ma, Ming Zhao, Yunfei Du. Impact of the strong Allee effect in a predator-prey model[J]. AIMS Mathematics, 2022, 7(9): 16296-16314. doi: 10.3934/math.2022890
In this work, we propose and investigate a new predator-prey model with strong Allee effect in prey and Holling type Ⅱ functional response in predator. By performing a comprehensive dynamical analysis, we first derive the existence and stability of all the possible equilibria of the system and the system undergoes two transcritical bifurcations and one Hopf-bifurcation. Next, we have calculated the first Lyapunov coefficient and find the Hopf-bifurcation in this model is supercritical and a stable limit cycle is born. Then, by comparing the properties of the system with and without Allee effect, we show that the strong Allee effect is of great importance to the dynamics. It can drive the system to instability. Specifically, Allee effect can increase the extinction risk of populations and has the ability to switch the system's stability to limit cycle oscillation from stable node. Moreover, numerical simulations are presented to prove the validity of our findings.

| [1] | N. Bacaer, A short history of mathematical population dynamics, Springer London, 2011. |
| [2] |
S. Bentout, A. Tridane, S. Djilali, T. M. Touaoula, Age-structured modeling of COVID-19 epidemic in the USA, UAE and Algeria, Alex. Eng. J., 60 (2021), 401–411. https://doi.org/10.1016/j.aej.2020.08.053 doi: 10.1016/j.aej.2020.08.053

|
| [3] | S. Djilali, S. Bentout, S. Kumar, T. M. Touaoula, Approximating the asymptomatic infectious cases of the COVID-19 disease in Algeria and India using a mathematical model, Int. J. Model. Simul. SC, 2250028 (2022). |
| [4] |
A. Singh, A. Parwaliya, A. Kumar, Hopf bifurcation and global stability of density-dependent model with discrete delays involving Beddington-DeAngelis functional response, Math. Method. Appl. Sci., 44 (2021), 8838–8861. https://doi.org/10.1002/mma.7311 doi: 10.1002/mma.7311
|
| [5] |
A. Singh, P. Malik, Bifurcations in a modified Leslie-Gower predator-prey discrete model with Michaelis-Menten prey harvesting, J. Appl. Math. Comput., 67 (2021), 143–174. https://doi.org/10.1007/s12190-020-01491-9 doi: 10.1007/s12190-020-01491-9
|
| [6] |
H. I. Freedman, Stability analysis of a predator-prey system with mutual interference and density-dependent death rates, Bull. Math. Biol., 41 (1979), 67–78. https://doi.org/10.1016/S0092-8240(79)80054-3 doi: 10.1016/S0092-8240(79)80054-3
|
| [7] |
D. P. Hu, H. J. Cao, Stability and bifurcation analysis in a predator-prey system with Michaelis-Menten type predator harvesting, Nonlinear Anal.-Real, 33 (2017), 58–82. https://doi.org/10.1016/j.nonrwa.2016.05.010 doi: 10.1016/j.nonrwa.2016.05.010
|
| [8] |
L. Li, W. C. Zhao, Deterministic and stochastic dynamics of a modified Leslie-Gower prey-predator system with simplified Holling-type Ⅳ scheme, Math. Biosci. Eng., 18 (2021), 2813–2831. https://doi.org/10.3934/mbe.2021143 doi: 10.3934/mbe.2021143
|
| [9] |
S. Djilali, S. Bentout, Spatiotemporal patterns in a diffusive predator-prey model with prey social behavior, Acta. Appl. Math., 169 (2020), 1–19. https://doi.org/10.1007/s10440-019-00291-z doi: 10.1007/s10440-019-00291-z
|
| [10] |
S. Djilali, S. Bentout, Pattern formations of a delayed diffusive predator-prey model with predator harvesting and prey social behavior, Math. Method. Appl. Sci., 44 (2021), 9128–9142. https://doi.org/10.1002/mma.7340 doi: 10.1002/mma.7340
|
| [11] |
M. J. Groom, Allee effects limit population viability of an annual plant, Am. Nat., 151 (1998), 487–496. https://doi.org/10.1086/286135 doi: 10.1086/286135
|
| [12] |
M. Kuussaari, I. Saccheri, M. Camara, I. Hanski, Allee effect and population dynamics in the Glanville fritillary butterfly, Oikos, 82 (1998), 384–392. https://doi.org/10.2307/3546980 doi: 10.2307/3546980
|
| [13] |
F. Courchamp, T. Clutton-Brock, B. Grenfell, Inverse density dependence and the Allee effect, Trends Ecol. Evol., 14 (1999), 405–410. https://doi.org/10.1016/S0169-5347(99)01683-3 doi: 10.1016/S0169-5347(99)01683-3
|
| [14] |
M. A. Mccarthy, The Allee effect, finding mates and theoretical models, Ecol. Model., 103 (1997), 99–102. https://doi.org/10.1016/S0304-3800(97)00104-X doi: 10.1016/S0304-3800(97)00104-X
|
| [15] | F. Courchamp, L. Berec, J. Gascoigne, Allee effects in ecology and conservation, Oxford University Press, London, 2008. https://doi.org/10.1093/acprof:oso/9780198570301.001.0001 |
| [16] |
M. Wang, M. Kot, Speeds of invasion in a model with strong or weak Allee effects, Math. Biosci., 171 (2001), 83–97. https://doi.org/10.1016/S0025-5564(01)00048-7 doi: 10.1016/S0025-5564(01)00048-7
|
| [17] |
J. F. Wang, J. P. Shi, J. J. Wei, Predator-prey system with strong Allee effect in prey, J. Math. Biol., 62 (2011), 291–331. https://doi.org/10.1007/s00285-010-0332-1 doi: 10.1007/s00285-010-0332-1
|
| [18] |
U. Kumar, P. S. Mandal, E. Venturino, Impact of Allee effect on an eco-epidemiological system, Ecol. Complex., 42 (2020), 100828. https://doi.org/10.1016/j.ecocom.2020.100828 doi: 10.1016/j.ecocom.2020.100828
|
| [19] |
G. Voorn, L. Hemerik, M. P. Boer, B. W. Kooi, Heteroclinic orbits indicate over exploitation in predator-prey systems with a strong Allee effect, Math. Biosci., 209 (2007), 451–469. https://doi.org/10.1016/j.mbs.2007.02.006 doi: 10.1016/j.mbs.2007.02.006
|
| [20] |
F. M. Hilker, M. Langlais, H. Malchow, The Allee effect and infectious diseases: Extinction, multistability, and the (dis)appearance of oscillations, Am. Nat., 173 (2009), 72–88. https://doi.org/10.1086/593357 doi: 10.1086/593357
|
| [21] |
S. Bentout, A. Chekroun, T. Kuniya, Parameter estimation and prediction for coronavirus disease outbreak 2019 (COVID-19) in Algeria, AIMS Public Health, 7 (2020), 306–318. https://doi.org/10.3934/publichealth.2020026 doi: 10.3934/publichealth.2020026
|
| [22] |
S. Bentout, T. M. Touaoula, Global analysis of an infection age model with a class of nonlinear incidence rates, J. Math. Anal. Appl., 434 (2016), 1211–1239. https://doi.org/10.1016/j.jmaa.2015.09.066 doi: 10.1016/j.jmaa.2015.09.066
|
| [23] |
S. Djilali, A. Mezouaghi, O. Belhamiti, Bifurcation analysis of a diffusive predator-prey model with schooling behaviour and cannibalism in prey, Int. J. Math. Model. Numer. Opt., 11 (2021), 209. https://doi.org/10.1504/IJMMNO.2021.116676 doi: 10.1504/IJMMNO.2021.116676
|
| [24] |
J. Zu, M. Mimura, The impact of Allee effect on a predator-prey system with Holling type Ⅱ functional response, Appl. Math. Comput., 217 (2010), 3542–3556. https://doi.org/10.1016/j.amc.2010.09.029 doi: 10.1016/j.amc.2010.09.029
|
| [25] |
C. S. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, Mem. Entomol. Soc. Can., 97 (1965), 1–60. https://doi.org/10.4039/entm9745fv doi: 10.4039/entm9745fv
|
| [26] |
S. Y. Tang, Y. N. Xiao, L. S. Chen, Integrated pest management models and their dynamical behaviour, Bull. Math. Biol., 67 (2005), 115–135. https://doi.org/10.1016/j.bulm.2004.06.005 doi: 10.1016/j.bulm.2004.06.005
|
| [27] |
J. Zu, W. D. Wang, B. Zu, Evolutionary dynamics of prey-predator systems with Holling type Ⅱ, Math. Biosci. Eng., 4 (2007), 221–237. https://doi.org/10.3934/mbe.2007.4.221 doi: 10.3934/mbe.2007.4.221
|
| [28] |
K. B. Sun, T. H. Zhang, Y. Tian, Dynamics analysis and control optimization of a pest management predator-prey model with an integrated control strategy, Appl. Math. Comput., 292 (2017), 253–271. https://doi.org/10.1016/j.amc.2016.07.046 doi: 10.1016/j.amc.2016.07.046
|
| [29] |
K. X. Wang, Influence of feedback controls on the global stability of a stochastic predator-prey model with Holling type Ⅱ response and infinite delays, Discrete. Cont. Dyn.-B, 25 (2020), 1699–1714. https://doi.org/10.3934/dcdsb.2019247 doi: 10.3934/dcdsb.2019247
|
| [30] |
X. Y. Wu, H. Zheng, S. C. Zhang, Dynamics of a non-autonomous predator-prey system with Hassell-Varley-Holling Ⅱ function response and mutual interference, AIMS Math., 6 (2021), 6033–6049. https://doi.org/10.3934/math.2021355 doi: 10.3934/math.2021355
|
| [31] | Z. F. Zhang, T. R. Ding, W. Z. Huang, Z. X. Dong, Qualitative theory of differential equation, Beijing, Beijing, 1997. |
| [32] | J. Sotomayor, Dynamical systems: Generic bifurcations of dynamical systems, Academic Press, New York, 1973,561–582. https://doi.org/10.1016/B978-0-12-550350-1.50047-3 |
| [33] | L. Perko, Differential equations and dynamical systems, Springer, New York, 2001. |






Figures(7)

Yudan Ma, Ming Zhao, Yunfei Du. Impact of the strong Allee effect in a predator-prey model[J]. AIMS Mathematics, 2022, 7(9): 16296-16314. doi: 10.3934/math.2022890







 DownLoad:
DownLoad: