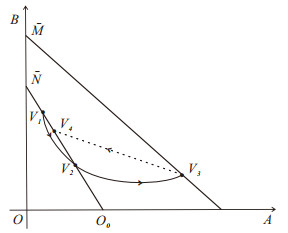
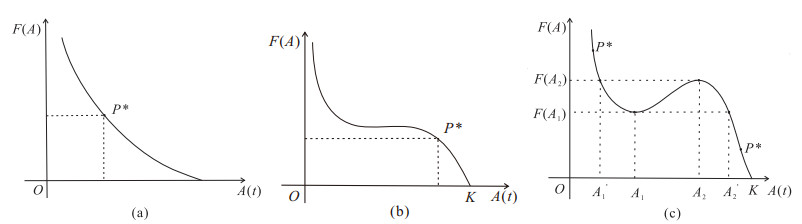
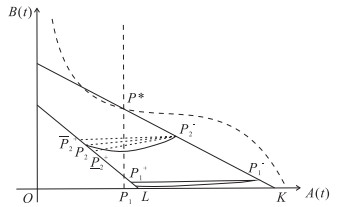
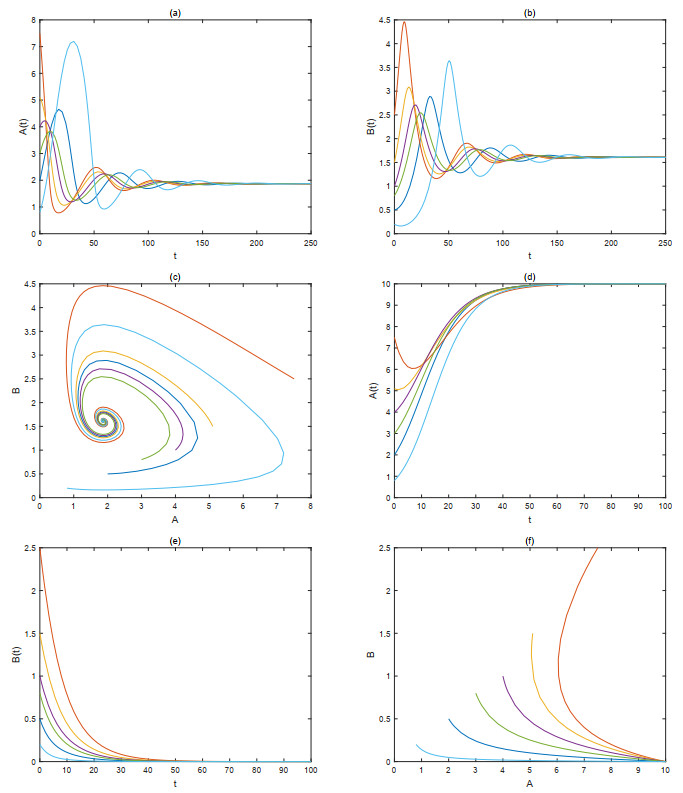
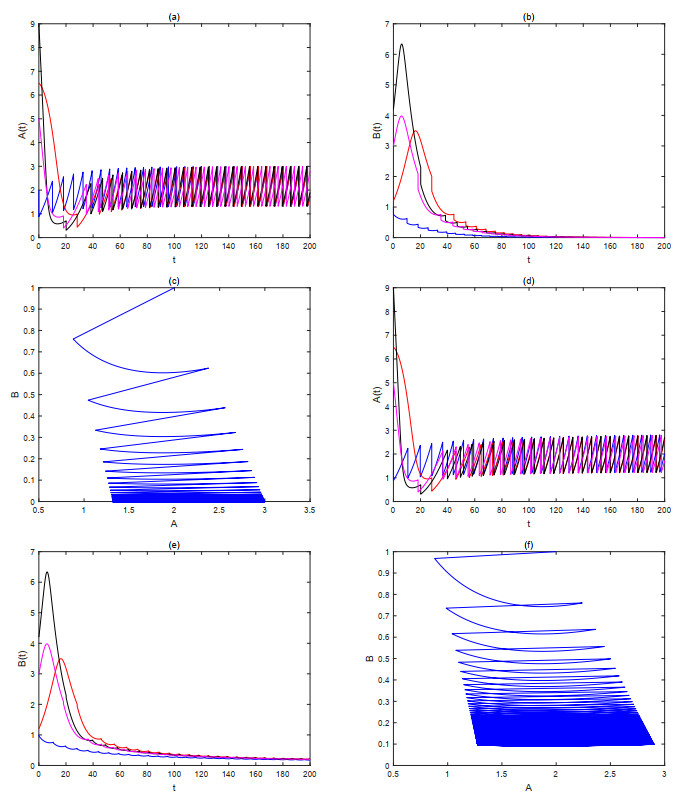
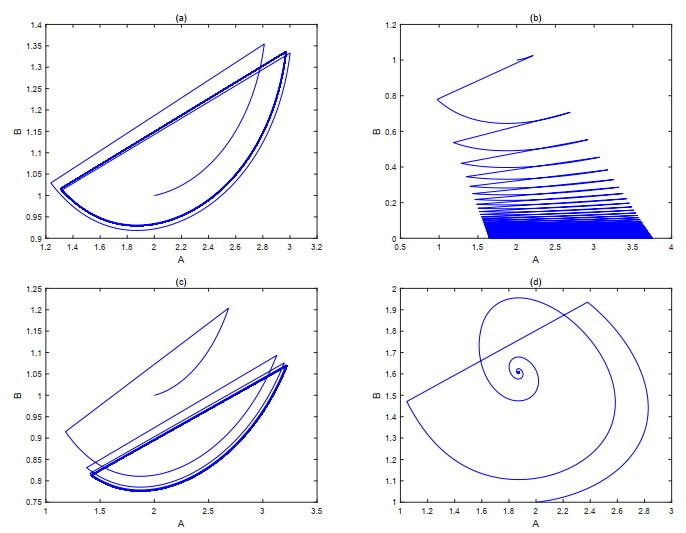
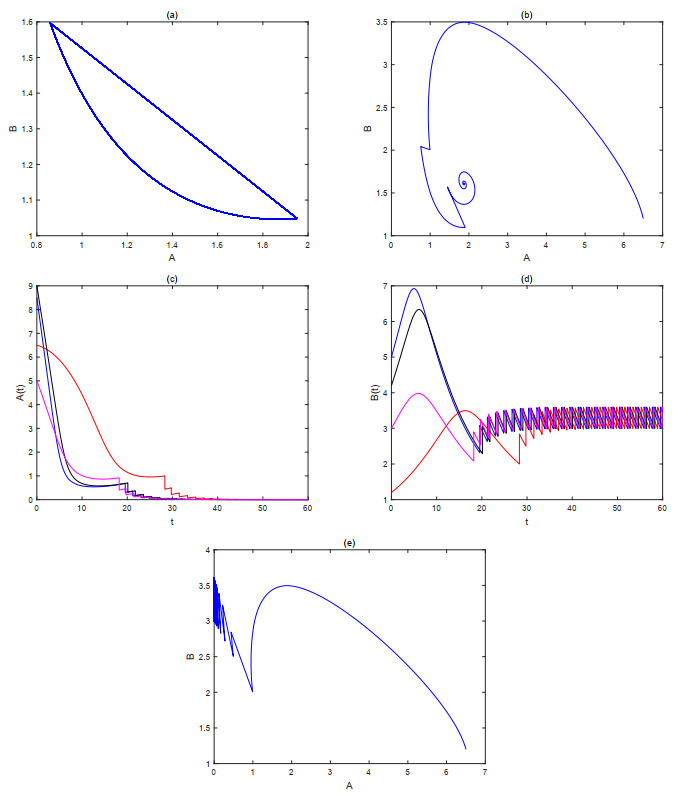
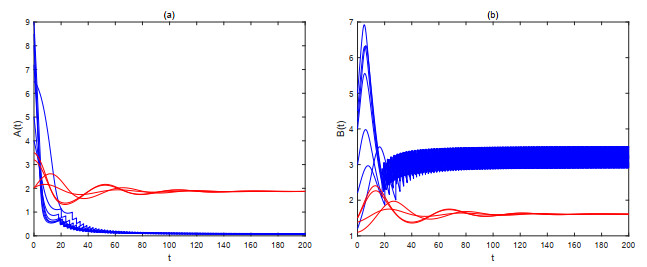
The current research presents a predator-prey model that incorporates both a Gilpin-Ayala growth function and a Holling type Ⅲ functional response. Two Lyapunov functions are established to confirm the global asymptotic stability of the positive equilibrium $ P^{*} $ and the predator extinction equilibrium $ P_{k} $. Considering ecological protection and commercial incentives, we also incorporated a weighted harvesting strategy and pulse control into the model. We investigated intricate dynamical problems instigated by the weighting harvesting and pulse effects, and affirmed the existence and local asymptotic stability of both predator-extinction periodic solution and positive order-1 periodic solution. In the end, a suite of numerical simulations were carried out using MATLAB, aiming to corroborate the theoretical findings and deliver conclusions rooted in a biological context.
Citation: Xiaohuan Yu, Mingzhan Huang. Dynamics of a Gilpin-Ayala predator-prey system with state feedback weighted harvest strategy[J]. AIMS Mathematics, 2023, 8(11): 26968-26990. doi: 10.3934/math.20231380
The current research presents a predator-prey model that incorporates both a Gilpin-Ayala growth function and a Holling type Ⅲ functional response. Two Lyapunov functions are established to confirm the global asymptotic stability of the positive equilibrium $ P^{*} $ and the predator extinction equilibrium $ P_{k} $. Considering ecological protection and commercial incentives, we also incorporated a weighted harvesting strategy and pulse control into the model. We investigated intricate dynamical problems instigated by the weighting harvesting and pulse effects, and affirmed the existence and local asymptotic stability of both predator-extinction periodic solution and positive order-1 periodic solution. In the end, a suite of numerical simulations were carried out using MATLAB, aiming to corroborate the theoretical findings and deliver conclusions rooted in a biological context.

| [1] |
F. Ayala, M. Gilpin, J. Ehrenfeld, Competition between species: theoretical models and experimental tests, Theor. Popul. Biol., 4 (1973), 331–356. https://doi.org/10.1016/0040-5809(73)90014-2 doi: 10.1016/0040-5809(73)90014-2

|
| [2] |
M. Amdouni, J. Alzabut, M. Samei, W. Sudsutad, C. Thaiprayoon, A generalized approach of the Gilpin-Ayala model with fractional derivatives under numerical simulation, Mathematics, 10 (2022), 3655. https://doi.org/10.3390/MATH10193655 doi: 10.3390/MATH10193655
|
| [3] |
S. Ai, J. Li, J. Yu, B. Zheng, Stage-structured models for interactive wild and periodically and impulsively released sterile mosquitoes, Discrete Cont. Dyn.-B, 27 (2022), 3039–3052. https://doi.org/10.3934/dcdsb.2021172 doi: 10.3934/dcdsb.2021172
|
| [4] |
I. Boubekri, H. Mazurek, A. Djebar, R. Amara, Harnessing Fishers' local knowledge and their perceptions: opportunities to improve management of coastal fishing in Mediterranean marine protected areas, J. Environ. Manage., 344 (2023), 118456. https://doi.org/10.1016/J.JENVMAN.2023.118456 doi: 10.1016/J.JENVMAN.2023.118456
|
| [5] |
J. Chen, J. Huang, S. Ruan, J. Wang, Bifurcations of invariant tori in predator-prey models with seasonal prey harvesting, SIAM J. Appl. Math., 73 (2013), 1876–1905. https://doi.org/10.1137/120895858 doi: 10.1137/120895858
|
| [6] | L. Chen, Pest control and geometric theory of semi-continuous dynamical system, J. Beihua Univ., 12 (2011), 9–11. |
| [7] |
M. Costa, E. Kaszkurewicz, A. Bhaya, L. Hsu, Achieving global convergence to an equilibrium population in predator-prey systems by the use of a discontinuous harvesting policy, Ecol. Model., 128 (2000), 89–99. https://doi.org/10.1016/S0304-3800(99)00220-3 doi: 10.1016/S0304-3800(99)00220-3
|
| [8] |
S. Debnath, P. Majumdar, S. Sarkar, U. Ghosh, Global dynamics of a prey-predator model with Holling type Ⅲ functional response in the presence of harvesting, J. Biol. Syst., 30 (2022), 225–260. https://doi.org/10.1142/S0218339022500073 doi: 10.1142/S0218339022500073
|
| [9] |
E. Gonz$\check{s}$lez-Olivares, A. Rojas-Palma, Multiple limit cycles in a gause predator-prey model with Holling Ⅲ functional response and Allee effect on prey, Bull. Math. Biol., 73 (2011), 1378–1397. https://doi.org/10.1007/s11538-010-9577-5 doi: 10.1007/s11538-010-9577-5
|
| [10] |
H. Guo, X. Song, L. Chen, Qualitative analysis of a korean pine forest model with impulse thinning measure, Appl. Math. Comput., 234 (2014), 203–213. https://doi.org/10.1016/j.amc.2014.02.034 doi: 10.1016/j.amc.2014.02.034
|
| [11] |
C. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, The Memoirs of the Entomological Society of Canada, 97 (1965), 5–60. https://doi.org/10.4039/entm9745fv doi: 10.4039/entm9745fv
|
| [12] |
M. Huang, S. Liu, X. Song, L. Chen, Periodic solutions and homoclinic bifurcation of a predator-prey system with two types of harvesting, Nonlinear Dyn., 73 (2013), 815–826. https://doi.org/10.1007/s11071-013-0834-7 doi: 10.1007/s11071-013-0834-7
|
| [13] |
M. Huang, X. Yu, S. Liu, X. Song, Dynamical behavior of a mosquito population suppression model composed of two sub-models, Int. J. Biomath., 16 (2023), 2250126. https://doi.org/10.1142/S1793524522501261 doi: 10.1142/S1793524522501261
|
| [14] |
M. Huang, X. Yu, S. Liu, Modeling and analysis of release strategies of sterile mosquitoes incorporating stage and sex structure of wild ones, Electron. Res. Arch., 31 (2023), 3895–3914. https://doi.org/10.3934/era.2023198 doi: 10.3934/era.2023198
|
| [15] |
M. Huang, X. Yu, Dynamic analysis of a mosquito population model with stage structure and periodic releases of sterile ones, AIMS Mathematics, 8 (2023), 18546–18565. https://doi.org/10.3934/math.2023943 doi: 10.3934/math.2023943
|
| [16] |
J. Jiao, L. Chen, S. Cai, Dynamical analysis of a biological resource management model with impulsive releasing and harvesting, Adv. Differ. Equ., 2012 (2012), 9. https://doi.org/10.1186/1687-1847-2012-9 doi: 10.1186/1687-1847-2012-9
|
| [17] |
Y. Jiang, Z. Liu, J. Yang, Y. Tan, Dynamics of a stochastic Gilpin-Ayala population model with Markovian switching and impulsive perturbations, Adv. Differ. Equ., 2020 (2020), 530. https://doi.org/10.1186/s13662-020-02900-w doi: 10.1186/s13662-020-02900-w
|
| [18] |
G. Kaniadakis, Novel predator-prey model admitting exact analytical solution, Phys. Rev. E, 106 (2022), 044401. https://doi.org/10.1103/PHYSREVE.106.044401 doi: 10.1103/PHYSREVE.106.044401
|
| [19] |
V. Křivan, The Lotka-Volterra predator-prey model with foraging-predation risk trade-offs, Am. Nat., 170 (2007), 771–782. https://doi.org/10.1086/522055 doi: 10.1086/522055
|
| [20] |
F. Rihan, H. Alsakaji, C. Rajivganthi, Stability and Hopf bifurcation of three-species prey-predator system with delays and Allee effect, Complexity, 2020 (2020), 7306412. https://doi.org/10.1155/2020/7306412 doi: 10.1155/2020/7306412
|
| [21] |
F. Souna, P. Tiwari, M. Belabbas, Y. Menacer, A predator-prey system with prey social behavior and generalized Holling Ⅲ functional response: role of predator-taxis on spatial patterns, Math. Method. Appl. Sci., 46 (2023), 13991–14006. https://doi.org/10.1002/MMA.9300 doi: 10.1002/MMA.9300
|
| [22] |
K. Sun, T. Zhang, Y. Tian, Theoretical study and control optimization of an integrated pest management predator-prey model with power growth rate, Math. Biosci., 279 (2016), 13–26. https://doi.org/10.1016/j.mbs.2016.06.006 doi: 10.1016/j.mbs.2016.06.006
|
| [23] |
S. Tang, B. Tang, A. Wang, Y. Xiao, Holling Ⅱ predator-prey impulsive semi-dynamic model with complex Poincar map, Nonlinear Dyn., 81 (2015), 1575–1596. https://doi.org/10.1007/s11071-015-2092-3 doi: 10.1007/s11071-015-2092-3
|
| [24] |
Y. Tian, Y. Gao, K. Sun, Global dynamics analysis of instantaneous harvest fishery model guided by weighted escapement strategy, Chaos Soliton. Fract., 164 (2022), 112597. https://doi.org/10.1016/j.chaos.2022.112597 doi: 10.1016/j.chaos.2022.112597
|
| [25] |
Y. Tian, Y. Gao, K. Sun, Qualitative analysis of exponential power rate fishery model and complex dynamics guided by a discontinuous weighted fishing strategy, Commun. Nonlinear Sci., 118 (2023), 107011. https://doi.org/10.1016/j.cnsns.2022.107011 doi: 10.1016/j.cnsns.2022.107011
|
| [26] |
M. Vasilova, Asymptotic behavior of a stochastic Gilpin-Ayala predator-prey system with time-dependent delay, Math. Comput. Model., 57 (2013), 764–781. https://doi.org/10.1016/j.mcm.2012.09.002 doi: 10.1016/j.mcm.2012.09.002
|
| [27] |
M. Vasilova, M. Jovanvic, Stochastic Gilpin-Ayala competition model with infinite delay, Appl. Math. Comput., 217 (2011), 4944–4959. https://doi.org/10.1016/j.amc.2010.11.043 doi: 10.1016/j.amc.2010.11.043
|
| [28] |
V. Volterra, Fluctuations in the abundance of a species considered mathematically, Nature, 118 (1926), 558–560. https://doi.org/10.1038/118558a0 doi: 10.1038/118558a0
|
| [29] |
W. Wei, W. Xu, J. Liu, Y. Song, S. Zhang, Stochastic bifurcation and Break-out of dynamic balance of predator-prey system with Markov switching, Appl. Math. Model., 117 (2023), 563–576. https://doi.org/10.1016/J.APM.2022.12.034 doi: 10.1016/J.APM.2022.12.034
|
| [30] |
H. Xu, T. Zhang, H. Cheng, Nonlinear control ecological model with complex discrete map, Commun. Nonlinear Sci., 118 (2023), 107019. https://doi.org/10.1016/j.cnsns.2022.107019 doi: 10.1016/j.cnsns.2022.107019
|
| [31] |
J. Xu, M. Huang, X. Song, Dynamics analysis of a two-species competitive system with state feedback impulsive control, Int. J. Biomath., 13 (2020), 2050007. https://doi.org/10.1142/S1793524520500072 doi: 10.1142/S1793524520500072
|
| [32] |
M. Zhang, L. Chen, Z. Li, Homoclinic bifurcation of a state feedback impulsive controlled prey-predator system with Holling-Ⅱ response, Nonlinear Dyn., 98 (2019), 929–942. https://doi.org/10.1007/s11071-019-05235-8 doi: 10.1007/s11071-019-05235-8
|
| [33] |
Q. Zhang, S. Tang, X. Zou, Rich dynamics of a predator-prey system with state-dependent impulsive controls switching between two means, J. Differ. Equations, 364 (2023), 336–377. https://doi.org/10.1016/J.JDE.2023.03.030 doi: 10.1016/J.JDE.2023.03.030
|
| [34] |
X. Zeng, L. Liu, W. Xie, Existence and uniqueness of the positive steady state solution for a Lotka-Volterra predator-prey model with a crowding term, Acta. Math. Sci., 40 (2020), 1961–1980. https://doi.org/10.1007/s10473-020-0622-7 doi: 10.1007/s10473-020-0622-7
|
Figures(8)

Xiaohuan Yu, Mingzhan Huang. Dynamics of a Gilpin-Ayala predator-prey system with state feedback weighted harvest strategy[J]. AIMS Mathematics, 2023, 8(11): 26968-26990. doi: 10.3934/math.20231380







 DownLoad:
DownLoad: