In this paper, we investigate a stochastic prey-predator model with Holling-type IV functional responses, anti-predatory behavior (referring to prey resistance to predator), gestation time delay of prey and Lévy noise. We investigate the existence and uniqueness of global positive solutions through Itô's formulation and Lyapunov's method. We also provide sufficient conditions for the persistence and extinction of prey-predator populations. Additionally, we examine the stability of the system distribution and validate our analytical findings through detailed numerical simulations. Our paper concludes with the implications of our results.
Citation: Chuanfu Chai, Yuanfu Shao, Yaping Wang. Analysis of a Holling-type IV stochastic prey-predator system with anti-predatory behavior and Lévy noise[J]. AIMS Mathematics, 2023, 8(9): 21033-21054. doi: 10.3934/math.20231071
In this paper, we investigate a stochastic prey-predator model with Holling-type IV functional responses, anti-predatory behavior (referring to prey resistance to predator), gestation time delay of prey and Lévy noise. We investigate the existence and uniqueness of global positive solutions through Itô's formulation and Lyapunov's method. We also provide sufficient conditions for the persistence and extinction of prey-predator populations. Additionally, we examine the stability of the system distribution and validate our analytical findings through detailed numerical simulations. Our paper concludes with the implications of our results.

| [1] | W. Thomas, Stability and complexity in model ecosystems, Princeton University Press, 1973. https://doi.org/10.1515/9780691206912 |
| [2] |
C. S. Holling, The functional response of predator to prey density and its role in mimicry and population regulation, Mem. Entomol. Soc. Can., 45 (1965), 1–60. https://doi.org/10.4039/entm9745fv doi: 10.4039/entm9745fv

|
| [3] |
W. W. Murdoch, A. Oaten, Predation and population stability, Adv. Ecol. Res., 9 (1975), 1–131. https://doi.org/10.1016/S0065-2504(08)60288-3 doi: 10.1016/S0065-2504(08)60288-3
|
| [4] |
L. A. Real, The kinetics of functional response, Am. Nat., 111 (1977), 289–300. https://doi.org/10.1086/283161 doi: 10.1086/283161
|
| [5] | N. V. Kampen, A. Heertjes, Statistical aspects of the predator-prey problem, J. Theor. Biol., 7 (1959), 1–36. |
| [6] |
Y. F. Shao, Fear and delay effects on a food chain system with two kinds of different functional responses, Int. J. Biomath., 34 (2023), 2350025. https://doi.org/10.1142/S1793524523500250 doi: 10.1142/S1793524523500250
|
| [7] |
S. Baba, A comprehensive cost-effectiveness analysis of control of maize streak virusdisease with Holling's Type II predation form and standard incidence, Results Phys., 40 (2022), 105862. https://doi.org/10.1016/j.rinp.2022.105862 doi: 10.1016/j.rinp.2022.105862
|
| [8] |
A. Singh, V. S. Sharma, Bifurcations and chaos control in a discrete-time prey-predator model with Holling type-II functional response and prey refuge, J. Comput. Appl. Math., 418 (2023), 114666. https://doi.org/10.1016/j.cam.2022.114666 doi: 10.1016/j.cam.2022.114666
|
| [9] |
C. S. Holling, Some characteristics of simple types of predation and parasitism, Can. Entomol., 91 (1959), 385–398. https://doi.org/10.4039/Ent91385-7 doi: 10.4039/Ent91385-7
|
| [10] |
F. Y. Wei, Uniform persistence of asymptotically periodic multispecies competition predator-prey systems with Holling III type functional response, Appl. Math. Comput., 170 (2005), 994–998. https://doi.org/10.1016/j.amc.2004.12.040 doi: 10.1016/j.amc.2004.12.040
|
| [11] |
Y. J. Huang, Stability analysis of a prey-predator model with holling type III response function incorporating a prey refuge, Appl. Math. Comput., 182 (2006), 672–683. https://doi.org/10.1016/j.amc.2006.04.030 doi: 10.1016/j.amc.2006.04.030
|
| [12] |
V. Madhusudanana, HOPF-bifurcation analysis of delayed computer virus modelwith holling type iii incidence function and treatment, Sci. Afr., 15 (2022), e01125. https://doi.org/10.1016/j.sciaf.2022.e01125 doi: 10.1016/j.sciaf.2022.e01125
|
| [13] |
W. Sokol, J. A. Howell, Kinetics of phenol exidation by washed cells, Biotechnol. Bioeng., 23 (1981), 2039–2049. https://doi.org/10.1002/bit.260230909 doi: 10.1002/bit.260230909
|
| [14] |
V. H. Edwards, The influence of high substrate concentrations on microbial kinetics, Biotechnol. Bioeng., 12 (1970), 679–712. https://doi.org/10.1002/bit.260120504 doi: 10.1002/bit.260120504
|
| [15] |
H. H. C. Alvino, M. Marvá, Group defense promotes coexistence in interference competition: The Holling type IV competitive response, Math. Comput. Simulat., 198 (2022), 426–445. https://doi.org/10.1016/j.matcom.2022.02.031 doi: 10.1016/j.matcom.2022.02.031
|
| [16] |
S. W. Zhang, A food chain model with impulsive perturbations and Holling IV functional response, Chaos Soliton. Fract., 26 (2005), 855–866. https://doi.org/10.1016/j.chaos.2005.01.053 doi: 10.1016/j.chaos.2005.01.053
|
| [17] |
S. W. Zhang, Chaos in periodically forced Holling type IV predator-prey system with impulsive perturbations, Chaos Soliton. Fract., 27 (2006), 980–990. https://doi.org/10.1016/j.chaos.2005.04.065 doi: 10.1016/j.chaos.2005.04.065
|
| [18] |
C. X. Shen, Permanence and global attractivity of the food-chain system with Holling IV type functional response, Appl. Math. Comput., 194 (2007), 179–185. https://doi.org/10.1016/j.amc.2007.04.019 doi: 10.1016/j.amc.2007.04.019
|
| [19] |
X. X. Liu, Q. D. Huang, The dynamics of a harvested predator-prey system with Holling type IV functional response, BioSystems, 169–170 (2018), 26–39. https://doi.org/10.1016/j.biosystems.2018.05.005 doi: 10.1016/j.biosystems.2018.05.005
|
| [20] |
K. Gopalsamy, Time lags and global stability in two species competition, Bull. Math. Biol., 42 (1980), 729–737. https://doi.org/10.1016/S0092-8240(80)80069-3 doi: 10.1016/S0092-8240(80)80069-3
|
| [21] |
Y. Zhao, S. L. Yuan, Q. M. Zhang, The effect of Lévy noise on the survival of a stochastic competitive model in an impulsive polluted environment, Appl. Math. Model., 40 (2016), 7583–7600. https://doi.org/10.1016/j.apm.2016.01.056 doi: 10.1016/j.apm.2016.01.056
|
| [22] |
Y. Zhao, S. L. Yuan, Stability in distribution of a stochastic hybrid competitive Lotka-Volterra model with Lévy jumps, Chaos Soliton. Fract., 85 (2016), 98–109. https://doi.org/10.1016/j.chaos.2016.01.015 doi: 10.1016/j.chaos.2016.01.015
|
| [23] |
C. Lu, Dynamical behavior of stochastic delay Lotka-Volterra competitive model with general Lévy jumps, Physica A, 531 (2019), 121730. https://doi.org/10.1016/j.physa.2019.121730 doi: 10.1016/j.physa.2019.121730
|
| [24] |
B. Tang, Y. Xiao, Bifurcation analysis of a predator-prey model with anti-predator behaviour, Chaos Soliton. Fract., 70 (2015), 58–68. https://doi.org/10.1016/j.chaos.2014.11.008 doi: 10.1016/j.chaos.2014.11.008
|
| [25] |
K. M. Comb, G. Shannon, S. M. Durant, Leadership in elephants: The adaptive value of age, P. Roy. Soc. B-Biol. Sci., 278 (2011), 3270–3276. https://doi.org/10.1098/rspb.2011.0168 doi: 10.1098/rspb.2011.0168
|
| [26] |
S. Creel, N. M. Creel, Limitation of African wild dogs by competition with larger carnivores, Conserv. Biol., 10 (1996), 526–538. https://doi.org/10.1046/j.1523-1739.1996.10020526.x doi: 10.1046/j.1523-1739.1996.10020526.x
|
| [27] | J. V. Craig, Effects of predation risk on reproductive behavior of northern fur seals, J. Mammal., 86 (2005), 1059–1067. |
| [28] | K. Gopalsamy, Stability and oscillations in delay differential equations of population dynamics, Springer Science+Business Media Dordrecht, 1992. https://doi.org/10.1007/978-94-015-7920-9 |
| [29] |
J. P. Tripathi, S. Abbas, M. Thakur, A density dependent delayed predator-prey model with Beddington-DeAngelis type function response incorporating a prey refuge, Commun. Nonlinear Sci. Numer. Simul., 22 (2015), 427–450. https://doi.org/10.1016/j.cnsns.2014.08.018 doi: 10.1016/j.cnsns.2014.08.018
|
| [30] |
A. Martin, S. Ruan, Predator-prey models with delay and prey harvesting, J. Math. Biol., 43 (2001), 247–267. https://doi.org/10.1007/s002850100095 doi: 10.1007/s002850100095
|
| [31] |
C. J. Xu, W. Zhang, C. Aouiti, Z. X. Liu, L. Y. Yao, Comparative exploration on bifurcation behavior for integer-order and fractional-order delayed BAM neural networks, Nonlinear Anal. Model. Control, 27 (2022), 1030–1053. https://doi.org/10.15388/namc.2022.27.28491 doi: 10.15388/namc.2022.27.28491
|
| [32] |
S. Ruan, Delay differential equations in single species dynamics, Delay Differ. Equat. Appl., 205 (2006), 477–517. https://dx.doi.org/10.1007/1-4020-3647-7_11 doi: 10.1007/1-4020-3647-7_11
|
| [33] | J. Barbalat, Systems d'equations differentielles d'osci d'oscillations nonlinéaires, Romanian J. Pure Appl. Math., 4 (1959), 267–270. |
| [34] |
M. Liu, C. Z. Bai, Optimal harvesting of a stochastic logistic model with time delay, J. Nonlinear Sci., 25 (2015), 277–289. https://doi.org/10.1007/s00332-014-9229-2 doi: 10.1007/s00332-014-9229-2
|
| [35] |
M. Liu, P. S. Mandal, Dynamical behavior of a one-prey two-predator model with random perturbations, Commun. Nonlinear Sci. Numer. Simulat., 28 (2015), 123–137. https://doi.org/10.1016/j.cnsns.2015.04.010 doi: 10.1016/j.cnsns.2015.04.010
|
| [36] |
Y. Guo, Stochastic regime switching SIR model driven by Lévy noise, Physica A, 497 (2017), 1–11. https://doi.org/10.1016/j.physa.2017.02.053 doi: 10.1016/j.physa.2017.02.053
|
| [37] |
H. Qiu, Optimal harvesting of a stochastic delay tri-trophic food-chain model with Lévy jumps, Physica A, 492 (2018), 1715–1728. https://doi.org/10.1016/j.physa.2017.11.092 doi: 10.1016/j.physa.2017.11.092
|
| [38] |
J. H. Bao, X. R. Mao, G. Yin, C. G. Yuan, Competitive Lotka-Volterra population dynamics with jumps, Nonlinear Anal., 74 (2011), 6601–6616. https://doi.org/10.1016/j.na.2011.06.043 doi: 10.1016/j.na.2011.06.043
|
| [39] |
J. Yu, Stationary distribution and ergodicity of a stochastic food-chain model with Lévy jumps, Physica A, 482 (2017), 14–28. https://doi.org/10.1016/j.physa.2017.04.067 doi: 10.1016/j.physa.2017.04.067
|
| [40] |
M. Deng, Stability of a stochastic delay commensalism model with Lévy jumps, Physica A, 527 (2019), 121061. https://doi.org/10.1016/j.physa.2019.121061 doi: 10.1016/j.physa.2019.121061
|
| [41] |
C. Liu, Q. L. Zhang, Y. K. Li, Dynamical behavior in a hybrid stochastic triple delayed prey predator bioeconomic system with Lévy jumps, J. Franklin I., 356 (2019), 592–628. https://doi.org/10.1016/j.jfranklin.2018.11.015 doi: 10.1016/j.jfranklin.2018.11.015
|
| [42] |
Y. L. Zhou, S. L. Yuan, D. L. Zhao, Threshold behavior of a stochastic SIS model with Lévy jumps, Appl. Math. Comput., 275 (2016), 255–267. https://doi.org/10.1016/j.amc.2015.11.077 doi: 10.1016/j.amc.2015.11.077
|
| [43] |
H. Kunita, Itô's stochastic calculus: Its surprising power for applications, Stoch. Process. Appl., 120 (2010), 622–652. https://doi.org/10.1016/j.spa.2010.01.013 doi: 10.1016/j.spa.2010.01.013
|
| [44] | R. X. Xue, Y. F. Shao, Analysis of a stochastic predator-prey system with fear effect and Lévy noise, Adv. Cont. Discr. Mod., 72 (2022). https://doi.org/10.1186/s13662-022-03749-x |
| [45] |
C. Liu, X. Y. Xun, Q. L. Zhang, Y. K. Li, Dynamical analysis and optimal control in a hybrid stochastic double delayed bioeconomic system with impulsive contaminants emission and Lévy jumps, Appl. Math. Comput., 352 (2019), 99–118. https://doi.org/10.1016/j.amc.2019.01.045 doi: 10.1016/j.amc.2019.01.045
|
| [46] |
Z. Ma, G. Cui, W. Wang, Persistence and extinction of a population in a polluted environment, Math. Biosci., 101 (1990), 75–97. https://doi.org/10.1016/0025-5564(90)90103-6 doi: 10.1016/0025-5564(90)90103-6
|
| [47] |
X. Y. Li, X. R. Mao, Population dynamical behavior of non-autonomous Lotka-Volterra competitive system with random perturbation, Discrete Cont. Dyn-S., 24 (2009), 523–545. https://doi.org/10.3934/dcds.2009.24.523 doi: 10.3934/dcds.2009.24.523
|
| [48] | M. Kot, Elements of mathematical biology, Cambridge University Press, 2001. https://doi.org/10.1017/CBO9780511608520 |
| [49] |
C. J. Xu, D. Mu, Y. L. Pan, C. Aouiti, L. Y. Yao, Exploring bifurcation in a fractional-order predator-prey system with mixed delays, J. Appl. Anal. Comput., 13 (2023), 1119–1136. https://doi.org/10.11948/20210313 doi: 10.11948/20210313
|
| [50] |
C. J. Xu, W. Zhang, C. Aouiti, Z. X. Liu, L. Y. Yao, Bifurcation insight for a fractional-order stage-structured predator-prey system incorporating mixed time delays, Math. Method. Appl. Sci., 46 (2023), 7489–7513. https://doi.org/10.1002/mma.9041 doi: 10.1002/mma.9041
|
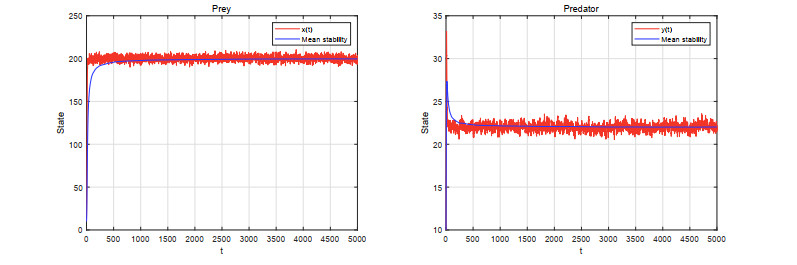
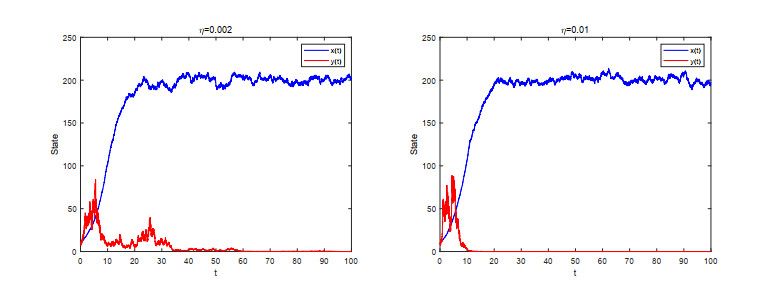
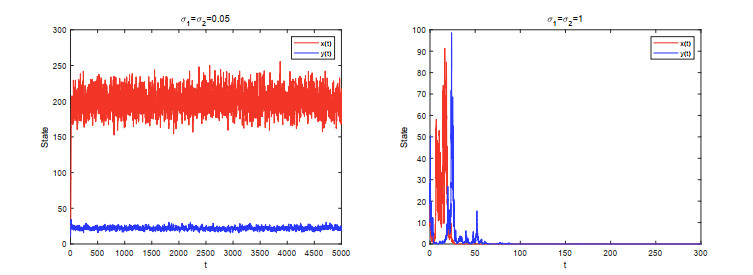
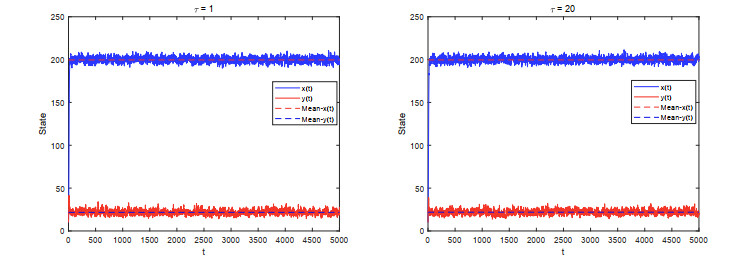
Figures(5)

Chuanfu Chai, Yuanfu Shao, Yaping Wang. Analysis of a Holling-type IV stochastic prey-predator system with anti-predatory behavior and Lévy noise[J]. AIMS Mathematics, 2023, 8(9): 21033-21054. doi: 10.3934/math.20231071







 DownLoad:
DownLoad: