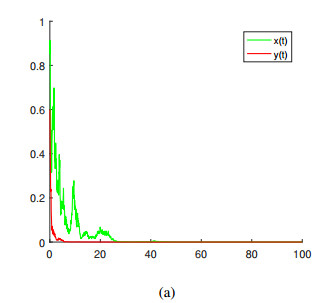
In this paper, a stochastic hybrid predator-prey model with Beddington-DeAngelis functional response and Lévy jumps is studied. Firstly, it is proved that the model has a unique global solution. Secondly, sufficient conditions for weak persistence in the mean and extinction of prey and predator populations are established. Finally, sufficient conditions for the existence and uniqueness of ergodic stationary distribution are established. Moreover, several numerical simulations are presented to illustrate the main results.
Citation: Hong Qiu, Yanzhang Huo, Tianhui Ma. Dynamical analysis of a stochastic hybrid predator-prey model with Beddington-DeAngelis functional response and Lévy jumps[J]. AIMS Mathematics, 2022, 7(8): 14492-14512. doi: 10.3934/math.2022799
In this paper, a stochastic hybrid predator-prey model with Beddington-DeAngelis functional response and Lévy jumps is studied. Firstly, it is proved that the model has a unique global solution. Secondly, sufficient conditions for weak persistence in the mean and extinction of prey and predator populations are established. Finally, sufficient conditions for the existence and uniqueness of ergodic stationary distribution are established. Moreover, several numerical simulations are presented to illustrate the main results.

| [1] |
M. Liu, C. X. Du, M. L. Deng, Persistence and extinction of a modified Leslie-Gower Holling-type II stochastic predator-prey model with impulsive toxicant input in polluted environments, Nonlinear Anal.-Hybri., 27 (2018), 177–190. http://doi.org/10.1016/j.nahs.2017.08.001 doi: 10.1016/j.nahs.2017.08.001

|
| [2] |
Z. J. Wang, M. L. Deng, M. Liu, Stationary distribution of a stochastic ratio-dependent predator-prey system with regime-switching, Chaos Soliton. Fract., 142 (2021), 110462. https://doi.org/10.1016/j.chaos.2020.110462 doi: 10.1016/j.chaos.2020.110462
|
| [3] | R. J. Swift, A stochastic predator-prey model, Bull. Irish Math. Soc., 48 (2002), 57–63. |
| [4] |
Q. Liu, D. Q. Jiang, T. Hayat, Dynamics of stochastic predator-prey models with distributed delay and stage structure for prey, Int. J. Biomath., 14 (2021), 2150020. https://doi.org/10.1142/S1793524521500200 doi: 10.1142/S1793524521500200
|
| [5] | S. Q. Zhang, T. H. Zhang, S. L. Yuan, Dynamics of a stochastic predator-prey model with habitat complexity and prey aggregation, Ecol. Complexity. 45 (2021), 100889. https://doi.org/10.1016/j.ecocom.2020.100889 |
| [6] |
L. L. Jia, Analysis for a delayed three-species predator-prey model with feedback controls and prey diffusion, J. Math., 2020 (2020), 5703859. https://doi.org/10.1155/2020/57038599 doi: 10.1155/2020/57038599
|
| [7] |
J. R. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Anim. Ecol., 44 (1975), 331–341. https://doi.org/10.2307/3866 doi: 10.2307/3866
|
| [8] |
D. L. DeAngelis, R. A. Goldsten, R. V. O'Neill, A model for trophic interaction, Ecology, 56 (1975), 881–892. https://doi.org/10.2307/1936298 doi: 10.2307/1936298
|
| [9] |
M. Liu, P. S. Mandal, Dynamical behavior of a one-prey two-predator model with random perturbations, Commun. Nonlinear Sci., 28 (2015), 123–137. https://doi.org/10.1016/j.cnsns.2015.04.010 doi: 10.1016/j.cnsns.2015.04.010
|
| [10] |
N. H. Du, V. H. Sam, Dynamics of a stochastic Lotka-Volterra model perturbed by white noise, J. Math. Anal. Appl., 324 (2006), 82–97. https://doi.org/10.1016/j.jmaa.2005.11.064 doi: 10.1016/j.jmaa.2005.11.064
|
| [11] |
D. Q. Jiang, N. Z. Shi, X. Y. Li, Global stability and stochastic permanence of a nonautonomous logistic equation with random perturbation, J. Math. Anal. Appl., 340 (2008), 588–597. https://doi.org/10.1016/j.jmaa.2007.08.014 doi: 10.1016/j.jmaa.2007.08.014
|
| [12] |
X. R. Mao, G. Marion, E. Renshaw, Environmental Brownian noise suppresses explosions in population dynamics, Stoch. Proc. Appl., 97 (2002), 95–110. https://doi.org/10.1016/S0304-4149(01)00126-0 doi: 10.1016/S0304-4149(01)00126-0
|
| [13] | X. R. Mao, C. G. Yuan, Stochastic differential equations with Markovian switching, London: Imperial College Press, 2006. https://doi.org/10.1142/p473 |
| [14] |
H. Wang, M. Liu, Stationary distribution of a stochastic hybrid phytoplankton-zooplankton model with toxin-producing phytoplankton, Appl. Math. Lett., 101 (2020), 106077. https://doi.org/10.1016/j.aml.2019.106077 doi: 10.1016/j.aml.2019.106077
|
| [15] |
D. G. Li, M. Liu, Invariant measure of a stochastic food-limited population model with regime switching, Math. Comput. Simulat., 178 (2020), 16–26. https://doi.org/10.1016/j.matcom.2020.06.003 doi: 10.1016/j.matcom.2020.06.003
|
| [16] |
C. Jeffries, Stability of predation ecosystem models, Ecology, 57 (1976), 1321–1325. https://doi.org/10.2307/1935058 doi: 10.2307/1935058
|
| [17] |
J. H. Bao, J. H. Shao, Permanence and extinction of regime-switching predator-prey models, SIAM J. Math. Anal., 48 (2016), 725–739. https://doi.org/10.1137/15M1024512 doi: 10.1137/15M1024512
|
| [18] |
C. Zhu, G. Yin, On competitive Lotka-Volterra model in random environments, J. Math. Anal. Appl., 357 (2009), 154–170. https://doi.org/10.1016/j.jmaa.2009.03.066 doi: 10.1016/j.jmaa.2009.03.066
|
| [19] |
M. Liu, Dynamics of a stochastic regime-switching predator-prey model with modified Leslie-Gower Holling-type II schemes and prey harvesting, Nonlinear Dyn., 96 (2019), 417–442. https://doi.org/10.1007/s11071-019-04797-x doi: 10.1007/s11071-019-04797-x
|
| [20] |
J. H. Bao, X. R. Mao, G. G. Yin, C. G. Yuan, Competitive Lotka-Volterra population dynamics with jumps, Nonlinear Anal.-Theor, 74 (2011), 6601–6616. https://doi.org/10.1016/j.na.2011.06.043 doi: 10.1016/j.na.2011.06.043
|
| [21] |
J. H. Bao, C. G. Yuan, Stochastic population dynamics driven by Lévy noise, J. Math. Anal. Appl., 391 (2012), 363–375. https://doi.org/10.1016/j.jmaa.2012.02.043 doi: 10.1016/j.jmaa.2012.02.043
|
| [22] | Y. C. Zang, J. P. Li, A dynamics of a stochastic predator-prey system with Beddington-DeAngelis functional response and Lévy jumps, Acta Math. Appl. Sin., 38 (2015), 340–349. |
| [23] |
H. Qiu, M. Liu, K. Wang, Y. Wang, Dynamics of a stochastic predator-prey system with Beddington-DeAngelis functional response, Appl. Math. Comput., 219 (2012), 2303–2312. https://doi.org/10.1016/j.amc.2012.08.077 doi: 10.1016/j.amc.2012.08.077
|
| [24] | V. Y. Krasin, A. V. Melnikov, On comparison theorem and its applications to finance, In: Optimality and risk modern trends in mathematical Finance, Berlin: Springer, 2009. https://doi.org/10.1007/978-3-642-02608-98 |
| [25] |
J. H. Bao, C. G. Yuan, Comparison theorem for stochastic differential delay equations with jumps, Acta Appl. Math., 116 (2011), 119. https://doi.org/10.1007/s10440-011-9633-7 doi: 10.1007/s10440-011-9633-7
|
| [26] |
X. L. Zou, K. Wang, Optimal harvesting for a stochastic regime-switching logistic diffusion system with jumps, Nonlinear Anal. Hybri., 13 (2014), 32–44. https://doi.org/10.1016/j.nahs.2014.01.001 doi: 10.1016/j.nahs.2014.01.001
|
| [27] |
M. Liu, Y. Zhu, Stationary distribution and ergodicity of a stochastic hybrid competition model with Lévy jumps, Nonlinear Anal. Hybri., 30 (2018), 225–239. https://doi.org/10.1016/j.nahs.2018.05.002 doi: 10.1016/j.nahs.2018.05.002
|
| [28] |
M. Liu, C. z. Bai, Dynamics of a stochastic one-prey two-predator model with Lévy jumps, Appl. Math. Comput., 284 (2016), 308–321. https://doi.org/10.1016/j.amc.2016.02.033 doi: 10.1016/j.amc.2016.02.033
|
| [29] |
S. Li, X. N. Zhang, Dynamics of a stochastic non-autonomous predator-prey system with Beddington-DeAngelis functional response, Adv. Differ. Equ., 2013 (2013), 19. https://doi.org/10.1186/1687-1847-2013-19 doi: 10.1186/1687-1847-2013-19
|
| [30] |
R. Lipster, A strong law of large numbers for local martingales, Stochastics, 3 (1980), 217–228. https://doi.org/10.1080/17442508008833146 doi: 10.1080/17442508008833146
|
| [31] |
J. L. Lv, K. Wang, M. Liu, Dynamical properties of a stochastic two-species Schoener's competitive model, Int. J. Biomath., 5 (2012), 1250035. https://doi.org/10.1142/S1793524511001751 doi: 10.1142/S1793524511001751
|
| [32] | I. Barbǎlat, Systemes d'equations differential d'oscillations nonlinearies (in Romanian), Revue des Math. Pures et Appl., 4 (1959), 267–270. |
| [33] | X. R. Mao, Stochastic differential equations and applications, 2 Eds., England: International Publishers in Science and Technology, 2007. |
| [34] | D. Applebaum, Lévy processes and stochastic calculus, 2 Eds., Cambridge: Cambridge Unversity Press, 2009. https://doi.org/10.1017/CBO9780511809781 |
| [35] |
G. B. Kallianpur, Stochastic differential equations and diffusion processes, Technometrics, 25 (1983), 208. https://doi.org/10.1080/00401706.1983.10487861 doi: 10.1080/00401706.1983.10487861
|
| [36] | G. Da Prato, J. Zabczyk, Ergodicity for infinite dimensional systems, Cambridge: Cambridge University Press, 1996. https://doi.org/10.1017/CBO9780511662829 |
| [37] |
E. S. Poloczanska, R. M. Cook, G. D. Ruxton, P. J. Wright, Fishing vs. natural recruitment variation in sandeels as a cause of seabird breeding failure at Shetland: A modelling approach, Ices J. Mar. Sci., 61 (2004), 788–797. https://doi.org/10.1016/j.icesjms.2004.03.030 doi: 10.1016/j.icesjms.2004.03.030
|
| [38] |
D. Gremillet, T. Boulinier, Spatial ecology and conservation of seabirds facing global climate change: a review, Mar. Ecol. Prog. Ser., 391 (2009), 121–137. https://doi.org/10.3354/meps08212 doi: 10.3354/meps08212
|
| [39] |
F. H. Vargas, R. C. Lacy, P. J. Johnson, A. Steinfurth, R. J. M. Crawford, P. D. Boersma, et al., Modelling the effect of El Nio on the persistence of small populations: The Galapagos penguin as a case study, Biol. Conserv., 137 (2007), 138–148. https://doi.org/10.1016/j.biocon.2007.02.005 doi: 10.1016/j.biocon.2007.02.005
|
| [40] |
B. H. Becker, M. Z. Peery, S. R. Beissinger, Ocean climate and prey availability affect the trophic level and reproductive success of the marbled murrelet, an endangered seabird, Mar. Ecol. Prog. Ser., 329 (2007), 267–279. https://doi.org/10.3354/meps329267 doi: 10.3354/meps329267
|





Figures(6) / Tables(1)

Hong Qiu, Yanzhang Huo, Tianhui Ma. Dynamical analysis of a stochastic hybrid predator-prey model with Beddington-DeAngelis functional response and Lévy jumps[J]. AIMS Mathematics, 2022, 7(8): 14492-14512. doi: 10.3934/math.2022799







 DownLoad:
DownLoad: