We study the dynamical properties of a discrete-time host-parasitoid model with Holling type III functional response. It is shown that flip bifurcation and Neimark-Sacker bifurcation occur in certain parameter regimes. A sufficient condition based on the model parameters for which both populations can coexist is derived. The boundedness, existence and local stability of the unique equilibrium are proved. In addition, the numerical simulations have been done, in addition to supporting the analytical findings, more behaviors are extracted from the model in a two-dimensional parameter space. Finally, we emphasize the importance of clearly presenting biological assumptions that are inherent to the structure of a discrete model.
Citation: Xijuan Liu, Yun Liu. Stability and bifurcation analysis of a discrete-time host-parasitoid model with Holling III functional response[J]. AIMS Mathematics, 2023, 8(10): 22675-22692. doi: 10.3934/math.20231154
We study the dynamical properties of a discrete-time host-parasitoid model with Holling type III functional response. It is shown that flip bifurcation and Neimark-Sacker bifurcation occur in certain parameter regimes. A sufficient condition based on the model parameters for which both populations can coexist is derived. The boundedness, existence and local stability of the unique equilibrium are proved. In addition, the numerical simulations have been done, in addition to supporting the analytical findings, more behaviors are extracted from the model in a two-dimensional parameter space. Finally, we emphasize the importance of clearly presenting biological assumptions that are inherent to the structure of a discrete model.

| [1] |
J. R. Beddington, C. A. Free, J. H. Lawton, Dynamics complexity in predator-prey models framed in difference equations, Nature, 255 (1975), 58–60. https://doi.org/10.1038/255058a0 doi: 10.1038/255058a0

|
| [2] |
Y. Xiao, S. Tang, The effect of initial density and parasitoid intergenerational survival rate on classical biological control, Chaos Soltion. Fract., 37 (2008), 1048–1058. https://doi.org/10.1016/j.chaos.2006.10.002 doi: 10.1016/j.chaos.2006.10.002
|
| [3] |
Q. Din, M. Hussain, Controlling chaos and Neimark-Sacker bifurcation in a host-parasitoid model, Asian J. Control, 21 (2019), 1202–1215. https://doi.org/10.1002/asjc.1809 doi: 10.1002/asjc.1809
|
| [4] |
A. Singh, B. Emerick, Generalized stability conditions for host-parasitoid population dynamics: Implications for biological control, Ecol. Model., 456 (2021), 109656. https://doi.org/10.1016/j.ecolmodel.2021.109656 doi: 10.1016/j.ecolmodel.2021.109656
|
| [5] |
C. Xiang, J. C. Huang, S. G. Ruan, D. Xiao, Bifurcation analysis in a host-parasitoid model with Holling II functional response, J. Differ. Equ., 268 (2020), 4618–4662. https://doi.org/10.1016/j.jde.2019.10.036 doi: 10.1016/j.jde.2019.10.036
|
| [6] |
T. Wang, Y. Zhang, Chemical control for host-parasitoid model within the parasitism season and its complex dynamics, Discrete Dyn. Nat. Soc., 2016 (2016), 3989625. http://doi.org/10.1155/2016/3989625 doi: 10.1155/2016/3989625
|
| [7] |
H. Liu, K. Zhang, Y. Wei, M. Ma, Dynamic complex and bifurcation analysis of a host-parasitoid model with Allee effect and Holling type III functional response, Adv. Differ. Equ., 2019 (2019), 507. https://doi.org/10.1186/s13662-019-2430-8 doi: 10.1186/s13662-019-2430-8
|
| [8] |
T. Cabello, M. Gamez, Z. Varga, An improvement of the Holling type III functional response in entomophagous species model, J. Biol. Syst., 15 (2007), 515–524. https://doi.org/10.1142/S0218339007002325 doi: 10.1142/S0218339007002325
|
| [9] | Q. Din, N. Saleem, M. S. Shabbir, A class of discrete predator-prey interaction with bifurcation analysis and chaos control, Math. Model. Nat. Phenom., 15 (2020), 60. |
| [10] |
L. Li, G. Q. Sun, Z. Jin, Bifurcation and chaos in an epidemic model with nonlinear incidence rates, Appl. Math. Comput., 216 (2010), 1226–1234. https://doi.org/10.1016/j.amc.2010.02.014 doi: 10.1016/j.amc.2010.02.014
|
| [11] |
Z. Dai, B. Du, Global dynamic analysis of periodic solution for discrete-time inertial neural networks with delays, AIMS Mathematics, 6 (2021), 3242–3256. https://doi.org/10.3934/math.2021194 doi: 10.3934/math.2021194
|
| [12] |
A. Singh, V. S. Sharma, Bifurcation and chaos control in a discrete-time prey-predator model with Holling type-II functional response and prey refuge, J. Comput. Appl. Math., 418 (2023), 114666. https://doi.org/10.1016/j.cam.2022.114666 doi: 10.1016/j.cam.2022.114666
|
| [13] |
S. Tang, L. Chen, Chaos in functional response host-parasitoid ecosystem models, Chaos Soltion. Fract., 13 (2002), 875–884. https://doi.org/10.1016/S0960-0779(01)00063-7 doi: 10.1016/S0960-0779(01)00063-7
|
| [14] | J. Carr, Application of center manifold theory, New York: Springer, 1981. https://doi.org/10.1007/978-1-4612-5929-9 |
| [15] | J. Gukenheimer, P. Holmes, Nonlinear oscillations, dynamicsl systems, and bifurcations of vector field, New York: Springer, 1983. https://doi.org/10.1007/978-1-4612-1140-2 |
| [16] |
X. Yang, Uniform persistence and periodic solutions for a discrete predator-prey system with delays, J. Math. Anal. Appl., 316 (2006), 161–177. https://doi.org/10.1016/j.jmaa.2005.04.036 doi: 10.1016/j.jmaa.2005.04.036
|
| [17] |
X. Liu, Y. Chu, Y. Liu, Bifurcation and chaos in a host-parasitoid model with a lower bound for the host, Adv. Differ. Equ., 2018 (2018), 31. https://doi.org/10.1186/s13662-018-1476-3 doi: 10.1186/s13662-018-1476-3
|
| [18] |
A. Tassaddiq, M. S. Shabbir, Q. Din, H. Naaz, Discretization, bifurcation, and control for a class of predator-prey interactions, Fractal Fract., 6 (2022), 31. https://doi.org/10.3390/fractalfract6010031 doi: 10.3390/fractalfract6010031
|
| [19] |
X. B. Rao, Y. D. Chu, Y. X. Chang, J. G. Zhang, Y. P. Tian Dynamics of a cracked rotor system with oil-film force in parameter space, Nonlinear Dyn., 88 (2017), 2347–2357. https://doi.org/10.1007/s11071-017-3381-9 doi: 10.1007/s11071-017-3381-9
|
| [20] |
X. Liu, P. Liu, Y. Liu, The existence of codimension-two bifurcations in a discrete-time SIR epidemic model, AIMS Mathematics, 7 (2021), 3360–3378. https://doi.org/10.3934/math.2022187 doi: 10.3934/math.2022187
|
| [21] |
P. C. Rech, The dynamics of a symmetric coupling of three modified quadratic maps, Chin. Phys. B., 22 (2013), 080202. https://doi.org/10.1088/1674-1056/22/8/080202 doi: 10.1088/1674-1056/22/8/080202
|
| [22] |
P. C. Rech, Organization of the periodicity in the parameter-space of a glycolysis discrete-time mathematical model, J. Math. Chem., 57 (2019), 632–637. https://doi.org/10.1007/s10910-018-0976-4 doi: 10.1007/s10910-018-0976-4
|
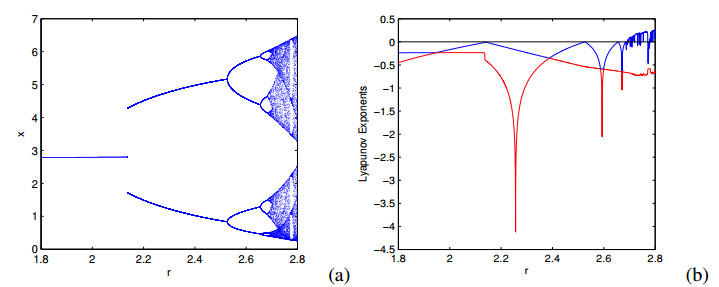





Figures(6)

Xijuan Liu, Yun Liu. Stability and bifurcation analysis of a discrete-time host-parasitoid model with Holling III functional response[J]. AIMS Mathematics, 2023, 8(10): 22675-22692. doi: 10.3934/math.20231154







 DownLoad:
DownLoad: