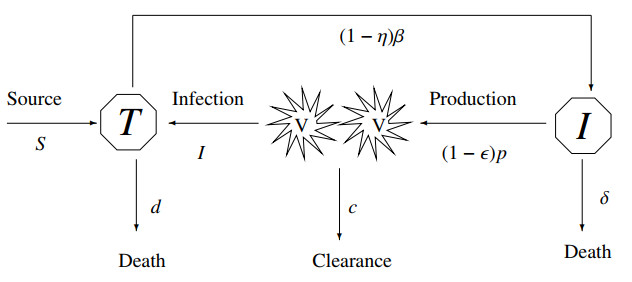
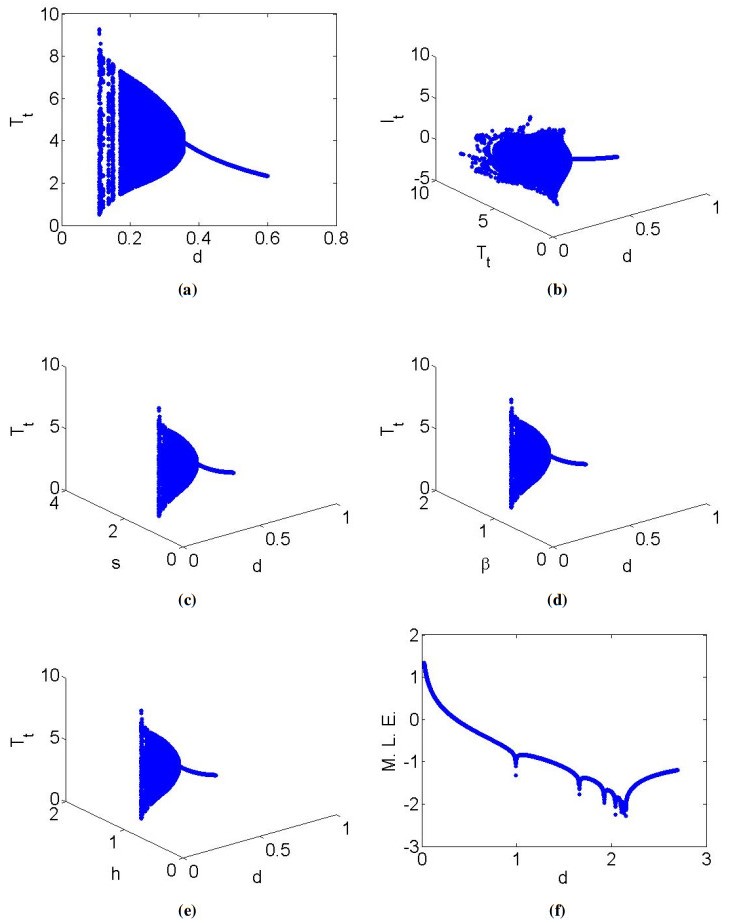
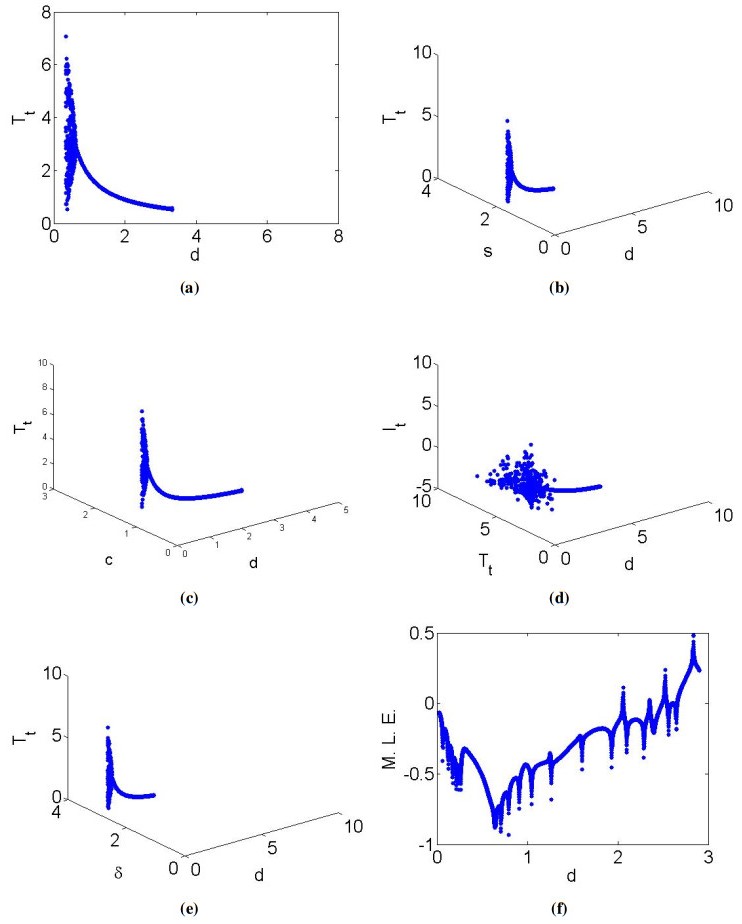
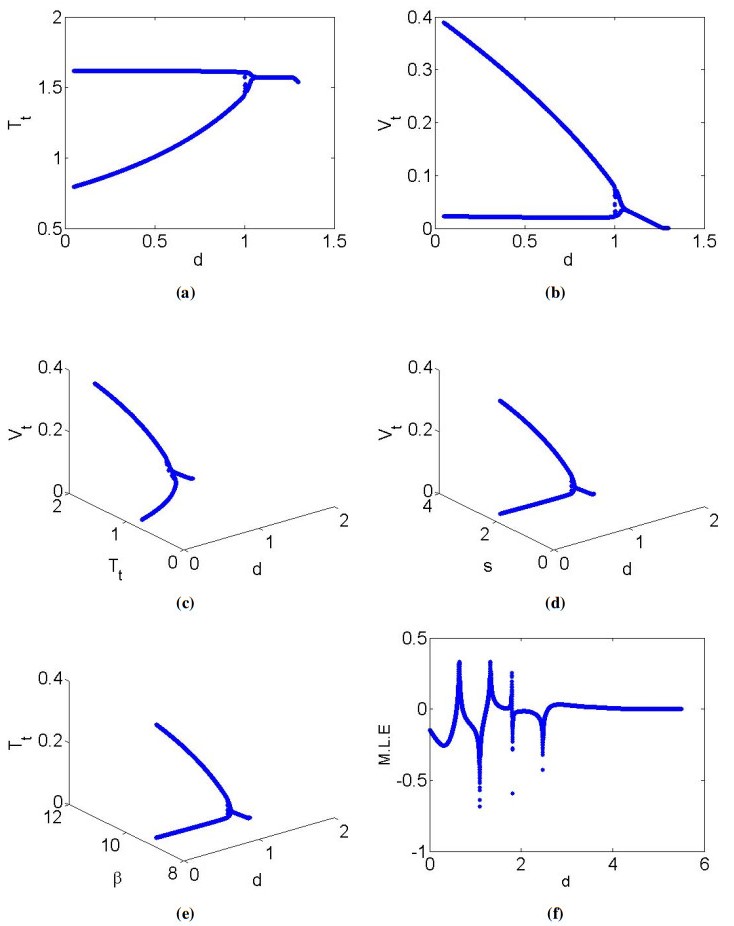
Mathematical models play a crucial role in understanding the dynamics of epidemic diseases by providing insights into how they spread and be controlled. In biomathematics, mathematical modeling is a powerful tool for interpreting the experimental results of biological phenomena related to disease transmission, offering precise and quantitative insights into the processes involved. This paper focused on a discrete mathematical model of the Hepatitis C virus (HCV) to analyze its dynamical behavior. Initially, we examined the local dynamics at steady states, providing a foundation for understanding the system's stability under various conditions. We then conducted a detailed bifurcation analysis, revealing that the discrete HCV model undergoes a Neimark-Sacker bifurcation at the uninfected steady state. Notably, our analysis showed that no period-doubling or fold bifurcations occur at this state. Further investigation at the infected steady state demonstrated the presence of both period-doubling and Neimark-Sacker bifurcations, which are characterized using explicit criteria. By employing a feedback control strategy, we explored chaotic behavior within the HCV model, highlighting the complex dynamics that can arise under certain conditions. Numerical simulations were conducted to verify the theoretical results, illustrating the model's validity and applicability. From a biological perspective, the insights gained from this analysis enhance our understanding of HCV transmission dynamics and potential intervention strategies. The presence of Neimark-Sacker bifurcation at the uninfected steady state implies that small perturbations could lead to oscillatory behavior, which may correspond to fluctuations in the number of infections over time. This finding suggests that maintaining stability at this steady state is critical for preventing outbreaks. The period-doubling and Neimark-Sacker bifurcations at the infected steady state indicate the potential for more complex oscillatory patterns, which could represent persistent cycles of infection and remission in a population. Finally, exploration of chaotic dynamics through feedback control highlights the challenges in predicting disease spread and the need for careful management strategies to avoid chaotic outbreaks.
Citation: Abdul Qadeer Khan, Ayesha Yaqoob, Ateq Alsaadi. Discrete Hepatitis C virus model with local dynamics, chaos and bifurcations[J]. AIMS Mathematics, 2024, 9(10): 28643-28670. doi: 10.3934/math.20241390
Mathematical models play a crucial role in understanding the dynamics of epidemic diseases by providing insights into how they spread and be controlled. In biomathematics, mathematical modeling is a powerful tool for interpreting the experimental results of biological phenomena related to disease transmission, offering precise and quantitative insights into the processes involved. This paper focused on a discrete mathematical model of the Hepatitis C virus (HCV) to analyze its dynamical behavior. Initially, we examined the local dynamics at steady states, providing a foundation for understanding the system's stability under various conditions. We then conducted a detailed bifurcation analysis, revealing that the discrete HCV model undergoes a Neimark-Sacker bifurcation at the uninfected steady state. Notably, our analysis showed that no period-doubling or fold bifurcations occur at this state. Further investigation at the infected steady state demonstrated the presence of both period-doubling and Neimark-Sacker bifurcations, which are characterized using explicit criteria. By employing a feedback control strategy, we explored chaotic behavior within the HCV model, highlighting the complex dynamics that can arise under certain conditions. Numerical simulations were conducted to verify the theoretical results, illustrating the model's validity and applicability. From a biological perspective, the insights gained from this analysis enhance our understanding of HCV transmission dynamics and potential intervention strategies. The presence of Neimark-Sacker bifurcation at the uninfected steady state implies that small perturbations could lead to oscillatory behavior, which may correspond to fluctuations in the number of infections over time. This finding suggests that maintaining stability at this steady state is critical for preventing outbreaks. The period-doubling and Neimark-Sacker bifurcations at the infected steady state indicate the potential for more complex oscillatory patterns, which could represent persistent cycles of infection and remission in a population. Finally, exploration of chaotic dynamics through feedback control highlights the challenges in predicting disease spread and the need for careful management strategies to avoid chaotic outbreaks.

| [1] |
M. Chong, M. Shahrill, L. Crossley, A. Madzvamuse, The stability analyses of the mathematical models of Hepatitis C virus infection, Mod. Appl. Sci., 9 (2015), 250–271. https://doi.org/10.5539/mas.v9n3p250 doi: 10.5539/mas.v9n3p250

|
| [2] |
J. Li, K. Men, Y. Yang, D. Li, Dynamical analysis on a chronic Hepatitis C virus infection model with immune response, J. Theoret. Biol., 365 (2015), 337–346. https://doi.org/10.1016/j.jtbi.2014.10.039 doi: 10.1016/j.jtbi.2014.10.039
|
| [3] |
F. A. Riha, A. A. Arafa, R. Rakkiyappan, C. Rajivganthi, Y. Xu, Fractional-order delay differential equations for the dynamics of Hepatitis C virus infection with IFN-$\alpha$ treatment, Alex. Eng. J., 60 (2021), 4761–4774. https://doi.org/10.1016/j.aej.2021.03.057 doi: 10.1016/j.aej.2021.03.057
|
| [4] |
A. Nangue, Global stability analysis of the original cellular model of Hepatitis C virus infection under therapy, Amer. J. Math. Comput. Model., 4 (2019), 58–65. https://doi.org/10.11648/j.ajmcm.20190403.12 doi: 10.11648/j.ajmcm.20190403.12
|
| [5] |
N. Ahmed, A. Raza, A. Akgül, Z. Iqbal, M. Rafiq, M. O. Ahmad, et al., New applications related to Hepatitis C model, AIMS Mathematics, 7 (2022), 11362–11381. https://doi.org/10.3934/math.2022634 doi: 10.3934/math.2022634
|
| [6] | J. M. Ntaganda, Modelling a therapeutic Hepatitis C virus dynamics, Int. J. Sci. Innov. Math. Res., 3 (2015), 1–10. |
| [7] |
G. Q. Sun, R. He, L. F. Hou, S. Gao, X. Luo, Q. Liu, et al., Dynamics of diseases spreading on networks in the forms of reaction-diffusion systems, Europhys. Lett. EPL, 147 (2024), 12001. https://doi.org/10.1209/0295-5075/ad5e1b doi: 10.1209/0295-5075/ad5e1b
|
| [8] |
O. RabieiMotlagh, L. Soleimani, Effect of mutations on stochastic dynamics of infectious diseases, a probability approach, Appl. Math. Comput., 451 (2023), 127993. https://doi.org/10.1016/j.amc.2023.127993 doi: 10.1016/j.amc.2023.127993
|
| [9] | R. M. Anderson, R. M. May, Infectious diseases of humans: Dynamics and control, Oxford university press, 1991. |
| [10] |
K. N. Nabi, C. N. Podder, Sensitivity analysis of chronic Hepatitis C virus infection with immune response and cell proliferation, Int. J. Biomath., 13 (2020), 2050017. https://doi.org/10.1142/S1793524520500175 doi: 10.1142/S1793524520500175
|
| [11] |
F. A. Rihan, M. Sheek-Hussein, A. Tridane, R. Yafia, Dynamics of Hepatitis C virus infection: Mathematical modeling and parameter estimation, Math. Model. Nat. Phenom., 12 (2017), 33–47. https://doi.org/10.1051/mmnp/201712503 doi: 10.1051/mmnp/201712503
|
| [12] |
M. Sadki, J. Danane, K. Allali, Hepatitis C virus fractional-order model: Mathematical analysis, Model. Earth Syst. Environ., 9 (2023), 1695–1707. https://doi.org/10.1007/s40808-022-01582-5 doi: 10.1007/s40808-022-01582-5
|
| [13] |
R. Shi, Y. Cui, Global analysis of a mathematical model for Hepatitis C virus transmissions, Virus Res., 217 (2016), 8–17. https://doi.org/10.1016/j.virusres.2016.02.006 doi: 10.1016/j.virusres.2016.02.006
|
| [14] |
M. Tahir, S. Inayat Ali Shah, G. Zaman, S. Muhammad, Ebola virus epidemic disease its modeling and stability analysis required abstain strategies, Cogent Biol., 4 (2018), 1488511. https://doi.org/10.1080/23312025.2018.1488511 doi: 10.1080/23312025.2018.1488511
|
| [15] |
P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [16] |
M. O. Onuorah, M. O. Nasir, M. S. Ojo, A. Ademu, A deterministic mathematical model for Ebola virus incorporating the vector population, Int. J. Math. Trends Technol., 30 (2016), 8–15. https://doi.org/10.14445/22315373/IJMTT-V30P502 doi: 10.14445/22315373/IJMTT-V30P502
|
| [17] |
M. Rafiq, W. Ahmad, M. Abbas, D. Baleanu, A reliable and competitive mathematical analysis of Ebola epidemic model, Adv. Differ. Equ., 2020 (2020), 540. https://doi.org/10.1186/s13662-020-02994-2 doi: 10.1186/s13662-020-02994-2
|
| [18] | E. Camouzis, G. Ladas, Dynamics of third-order rational difference equations with open problems and conjectures, New York: Chapman and Hall/CRC, 2007. https://doi.org/10.1201/9781584887669 |
| [19] | E. A. Grove, G. Ladas, Periodicities in nonlinear difference equations, New York: Chapman and Hall/CRC, 2004. https://doi.org/10.1201/9781420037722 |
| [20] | V. L. Kocic, G. Ladas, Global behavior of nonlinear difference equations of higher-order with applications, Dordrecht: Springer, 1993. https://doi.org/10.1007/978-94-017-1703-8 |
| [21] | H. Sedaghat, Nonlinear difference equations: Theory with applications to social science models, Dordrecht: Springer, 2003. https://doi.org/10.1007/978-94-017-0417-5 |
| [22] | M. R. S. Kulenović, G. Ladas, Dynamics of second-order rational difference equations: With open problems and conjectures, New York: Chapman and Hall/CRC, 2001. https://doi.org/10.1201/9781420035384 |
| [23] | A. Wikan, Discrete dynamical systems–with an introduction to discrete optimization problems, 2013. |
| [24] | J. Guckenheimer, P. Holmes, Nonlinear oscillations, dynamical systems, and bifurcations of vector fields, New York: Springer, 2013. https://doi.org/10.1007/978-1-4612-1140-2 |
| [25] | Y. A. Kuznetsov, Elements of applied bifurcation theorey, New York: Springer, 2004. https://doi.org/10.1007/978-1-4757-3978-7 |
| [26] |
H. N. Agiza, E. M. Elabbssy, H. El-Metwally, A. A. Elsadany, Chaotic dynamics of a discrete prey-predator model with Holling type Ⅱ, Nonlinear Anal. Real World Appl., 10 (2009), 116–129. https://doi.org/10.1016/j.nonrwa.2007.08.029 doi: 10.1016/j.nonrwa.2007.08.029
|
| [27] |
A. M. Yousef, S. M. Salman, A. A. Elsadany, Stability and bifurcation analysis of a delayed discrete predator-prey model, Int. J. Bifur. Chaos, 28 (2018), 1850116. https://doi.org/10.1142/S021812741850116X doi: 10.1142/S021812741850116X
|
| [28] |
A. Q. Khan, J. Ma, D. Xiao, Bifurcations of a two-dimensional discrete time plant-herbivore system, Commun. Nonlinear Sci. Numer. Simul., 39 (2016), 185–198. https://doi.org/10.1016/j.cnsns.2016.02.037 doi: 10.1016/j.cnsns.2016.02.037
|
| [29] |
A. Q. Khan, J. Ma, D. Xiao, Global dynamics and bifurcation analysis of a host-parasitoid model with strong Allee effect, J. Biol. Dyn., 11 (2017), 121–146. https://doi.org/10.1080/17513758.2016.1254287 doi: 10.1080/17513758.2016.1254287
|
| [30] | E. M. Elabbasy, H. N Agiza, H. El-Metwally, A. A. Elsadany, Bifurcation analysis, chaos and control in the Burgers mapping, Int. J. Nonlinear Sci., 4 (2007), 171–185. |
| [31] |
G. Wen, Criterion to identify hopf bifurcations in maps of arbitrary dimension, Phys. Rev. E, 72 (2005), 026201. https://doi.org/10.1103/PhysRevE.72.026201 doi: 10.1103/PhysRevE.72.026201
|
| [32] |
S. Yao, New bifurcation critical criterion of Flip-Neimark-Sacker bifurcations for two-parameterized family of $n$-dimensional discrete systems, Discrete Dyn. Nat. Soc., 2012 (2012), 264526. https://doi.org/10.1155/2012/264526 doi: 10.1155/2012/264526
|
| [33] |
B. Xin, T. Chen, J. Ma, Neimark-Sacker bifurcation in a discrete-time financial system, Discrete Dyn. Nat. Soc., 2010 (2010), 405639. https://doi.org/10.1155/2010/405639 doi: 10.1155/2010/405639
|
| [34] |
G. Wen, S. Chen, Q. Jin, A new criterion of period-doubling bifurcation in maps and its application to an inertial impact shaker, J. Sound Vib., 311 (2008), 212–223. https://doi.org/10.1016/j.jsv.2007.09.003 doi: 10.1016/j.jsv.2007.09.003
|
| [35] | S. Elaydi, An introduction to difference equations, New York: Springer, 1999. https://doi.org/10.1007/0-387-27602-5 |
| [36] | S. Lynch, Dynamical systems with applications using mathematica, Boston: Birkhäuser, 2007. https://doi.org/10.1007/978-0-8176-4586-1 |
Figures(5)

Abdul Qadeer Khan, Ayesha Yaqoob, Ateq Alsaadi. Discrete Hepatitis C virus model with local dynamics, chaos and bifurcations[J]. AIMS Mathematics, 2024, 9(10): 28643-28670. doi: 10.3934/math.20241390







 DownLoad:
DownLoad: