Antibiotic resistance is one of the top 10 public health problems that most affects humanity. In recent decades, plasmid-mediated antibiotic resistance (PMAR) has increased. However, due to the lack of knowledge about the biology of plasmids, there are gaps in the role played by them within antibiotic resistance. In this sense, properties that agree with the biological phenomenon and have contributed to the understanding of PMAR have been discovered from mathematical modeling. In this work, we focus on the role that the plasmid replication rate plays in the elimination or spread of bacteria both sensitive and resistant to antibiotics. Qualitative analysis reveals the existence of a free-bacteria equilibrium point, a resistant equilibrium point, and two coexistence equilibrium points (high and low bacterial load). If each bacterium (sensitive or resistant) produces at most one new bacterium, the infection will be controlled or eliminated. If each bacterium (sensitive or resistant) produces more than one bacteria, several scenarios of bacterial progression are presented that depend on the plasmid replication rate. The results suggest that plasmid replication is essential for the outcome of bacterial infection at the local level.
Citation: Eduardo Ibargüen-Mondragón, Miller Cerón Gómez, Edith M. Burbano-Rosero. Assessing the role of bacterial plasmid replication in a competition model of sensitive and resistant bacteria to antibiotics[J]. AIMS Mathematics, 2021, 6(9): 9446-9467. doi: 10.3934/math.2021549
Antibiotic resistance is one of the top 10 public health problems that most affects humanity. In recent decades, plasmid-mediated antibiotic resistance (PMAR) has increased. However, due to the lack of knowledge about the biology of plasmids, there are gaps in the role played by them within antibiotic resistance. In this sense, properties that agree with the biological phenomenon and have contributed to the understanding of PMAR have been discovered from mathematical modeling. In this work, we focus on the role that the plasmid replication rate plays in the elimination or spread of bacteria both sensitive and resistant to antibiotics. Qualitative analysis reveals the existence of a free-bacteria equilibrium point, a resistant equilibrium point, and two coexistence equilibrium points (high and low bacterial load). If each bacterium (sensitive or resistant) produces at most one new bacterium, the infection will be controlled or eliminated. If each bacterium (sensitive or resistant) produces more than one bacteria, several scenarios of bacterial progression are presented that depend on the plasmid replication rate. The results suggest that plasmid replication is essential for the outcome of bacterial infection at the local level.

| [1] | H. Lodish, A. Berk, P. Matsudaira, C. A. Kaiser, M. Krieger, M. P. Scott, et al., Biología Celular y Molecular, Editorial Médica Panamericana, Bogotá, 2016. |
| [2] |
L. A. Actis, M. E. Tolmasky, J. H. Crosa, Bacterial plasmids: Replication of extrachromosomal genetic elements encoding resistance to antimicrobial compounds, Front. Biosci., 4 (1999), 43–62. doi: 10.2741/Actis

|
| [3] |
R. X. Wang, J. Y. Wang, Y. C. Sun, B. L. Yang, A. L. Wang, Antibiotic resistance monitoring in Vibrio spp. isolated from rearing environment and intestines of abalone Haliotis diversicolor, Mar. Pollut. Bull., 101 (2015), 701–706. doi: 10.1016/j.marpolbul.2015.10.027
|
| [4] |
M. Zhou, Y. Xu, P. Ouyang, J. Ling, Q. Cai, L. Huang, et al., Evolution and distribution of resistance genes and bacterial community in water and biofilm of a simulated fish-duck integrated pond with stress, Chemosphere, 245 (2020), 125549. doi: 10.1016/j.chemosphere.2019.125549
|
| [5] | C. Schultsz, S. Geerlings, Plasmid-mediated resistance in Enterobacteriaceae, Drugs, 72 (2012), 1–16. |
| [6] | M. S. Ramirez, G. M. Traglia, D. L. Lin, T. Tran, M. E. Tolmasky, Plasmid‐mediated antibiotic resistance and virulence in gram‐negatives: The Klebsiella pneumoniae paradigm, Microbiol. Spectr., 2 (2014), 1–15. |
| [7] |
B. Li, F. Yin, X. Zhao, Y. Guo, W. Wang, P. Wang, et al., Colistin resistance gene mcr-1 mediates cell permeability and resistance to hydrophobic antibiotics, Front. Microbiol., 10 (2020), 1–7. doi: 10.3389/fcimb.2020.00001
|
| [8] |
L. G. Burman, R-plasmid transfer and its response to nalidixic acid, J. Bacteriol., 131 (1977), 76–81. doi: 10.1128/jb.131.1.76-81.1977
|
| [9] | E. A. Yurtsev, H. X. Chao, M. S. Datta, T. Artemova, J. Gore, Bacterial cheating drives the population dynamics of cooperative antibiotic resistance plasmids, Mol. Syst. Biol., 9 (2013), 1–7. |
| [10] |
D. Godwin, J. H. Slater, The influence of the growth environment on the stability of a drug resistance plasmid in Escherichia coli K12, J. Gen. Microbiol., 111 (1979), 201–210. doi: 10.1099/00221287-111-1-201
|
| [11] |
B. Daşbaşı, İ. Öztürk, Mathematical modelling of bacterial resistance to multiple antibiotics and immune system response, Springerplus, 5 (2016), 1–17. doi: 10.1186/s40064-015-1659-2
|
| [12] | M. Merdan, Z. Bekiryazici, T. Kesemen, T. Khaniyev, Comparison of stochastic and random models for bacterial resistance, Adv. Differ. Equations, 133 (2017), 1–19. |
| [13] | Q. J. Leclerc, J. A. Lindsay, G. M. Knight, Mathematical modelling to study the horizontal transfer of antimicrobial resistance genes in bacteria: Current state of the field and recommendations, J. R. Soc. Interface., 16 (2018), 1–10. |
| [14] |
J. N. Tetteh, F. Matthüs, E. A. Hernandez-Vargas, A survey of within-host and between-hosts modelling for antibiotic resistance, Biosystems, 196 (2020), 1–13. doi: 10.1016/j.biosystemseng.2020.05.010
|
| [15] |
E. Ibargüen-Mondragón, J. P. Romero-Leiton, L. Esteva, E. M. Burbano-Rosero, Mathematical modeling of bacterial resistance to antibiotics by mutations and plasmids, J. Biol. Syst., 24 (2016), 129–146. doi: 10.1142/S0218339016500078
|
| [16] |
E. Ibarguen-Mondragon, J. P. Romero-Leiton, L. Esteva, M. C. Gomez, S. P. Hidalgo-Bonilla, Stability and periodic solutions for a model of bacterial resistance to antibiotics caused by mutations and plasmids, Appl. Math. Model., 76 (2019), 238–251. doi: 10.1016/j.apm.2019.06.017
|
| [17] |
C. S. Hayes, D. A. Low, Signals of growth regulation in bacteria, Curr. Opin. Microbiol., 12 (2009), 667–673. doi: 10.1016/j.mib.2009.09.006
|
| [18] | K. N. Timmis, M. I. Gonzalez-Carrero, T. Sekizaki, F. Rojo, Biological activities specified by antibiotic resistance plasmids, J. Antimicrob. Chemother., 18 (1986), 1–12. |
| [19] | T. Wein, F. T. Stücker, N. F. Hülter, T. Dagan, Quantification of plasmid-mediated antibiotic resistance in an experimental evolution approach, J. Vis. Exp., 154 (2019), 1–7. |
| [20] | E. Ibarguen-Mondragon, K. Prieto, S. P. Hidalgo-Bonilla, A model on bacterial resistance considering a generalized law of mass action for plasmid replication, J. Biol. Syst., 28 (2021), 1–38. |
| [21] |
E. Ibargüen-Mondragón, L. Esteva, J. P. Romero-Leiton, S. P. Hidalgo-Bonilla, E. M. Burbano-Rosero, M. Cerón, et al., Mathematical modeling on bacterial resistance to multiple antibiotics caused by spontaneus mutations, Biosystems, 117 (2014), 60–67. doi: 10.1016/j.biosystems.2014.01.005
|
| [22] | J. P. Romero-Leiton, E. Ibargüen-Mondragón, Sobre la resistencia bacteriana a antibióticos de acción bactericida y bacteriostática, Rev. Integración, 32 (2014), 101–116. |
| [23] |
K. Nordström, S. Molin, J. Light, Control of replication of bacterial plasmids: Genetics, molecular biology, and physiology of the plasmid R1 system, Plasmid, 12 (1984), 71–90. doi: 10.1016/0147-619X(84)90054-4
|
| [24] | L. Perko, Differential Equations and Dynamical Systems, 2 Eds, New Yorks: Springer Science $ & $ Business Media, 2013. |
| [25] |
W. Karpińska, On bounded solutions of nonlinear first and second order equations with a Carathéodory function, J. Math. Anal. Appl., 334 (2007), 1462–1480. doi: 10.1016/j.jmaa.2007.01.037
|
| [26] |
D. S. Schmidt, Hopf's bifurcation theorem and the center theorem of Liapunov with resonance cases, J. Math. Anal. Appl., 63 (1978), 354–370. doi: 10.1016/0022-247X(78)90081-1
|
| [27] | M. W. Hisrch, S. Smale, Differential Equations, Dynamical Systems and Linear Algebra, 2 Eds, New York: Academic Press, 1974. |
| [28] |
V. N. Satyagal, P. Agrawal, A generalized model of plasmid replication, Biotechnol. Bioeng., 33 (1989), 1135–1144. doi: 10.1002/bit.260330909
|
| [29] | T. M. Apostol, Mathematical Analysis, Massachusetts: Addison-Wesley, 1960. |
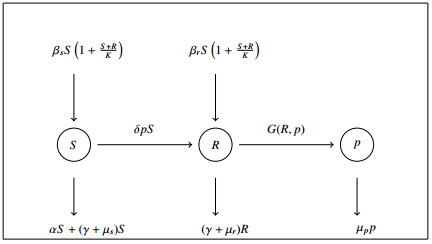
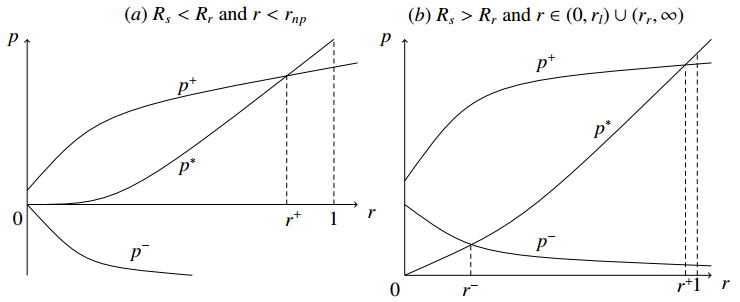
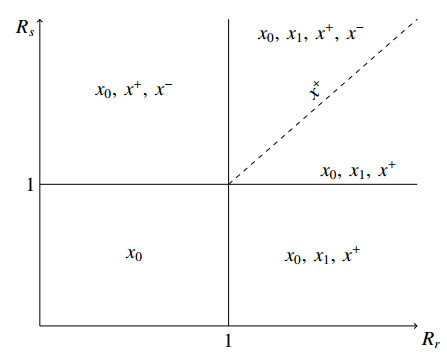
Figures(3)

Eduardo Ibargüen-Mondragón, Miller Cerón Gómez, Edith M. Burbano-Rosero. Assessing the role of bacterial plasmid replication in a competition model of sensitive and resistant bacteria to antibiotics[J]. AIMS Mathematics, 2021, 6(9): 9446-9467. doi: 10.3934/math.2021549







 DownLoad:
DownLoad: