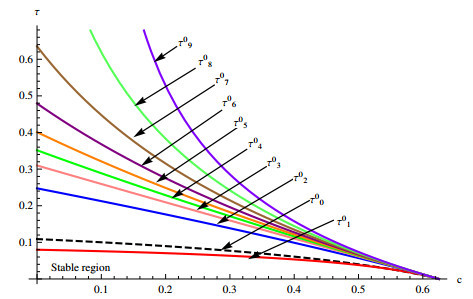
A delayed diffusive predator-prey system with nonlocal competition and generalist predators is considered. The local stability of the positive equilibrium and Hopf bifurcation at positive equilibrium is studied by using time delay as a parameter. In addition, the property of Hopf bifurcation is analyzed using the center manifold theorem and normal form method. It is determined that time delays can affect the stability of the positive equilibrium and induce spatial inhomogeneous periodic oscillation of prey and predator population densities.
Citation: Chenxuan Nie, Dan Jin, Ruizhi Yang. Hopf bifurcation analysis in a delayed diffusive predator-prey system with nonlocal competition and generalist predator[J]. AIMS Mathematics, 2022, 7(7): 13344-13360. doi: 10.3934/math.2022737
A delayed diffusive predator-prey system with nonlocal competition and generalist predators is considered. The local stability of the positive equilibrium and Hopf bifurcation at positive equilibrium is studied by using time delay as a parameter. In addition, the property of Hopf bifurcation is analyzed using the center manifold theorem and normal form method. It is determined that time delays can affect the stability of the positive equilibrium and induce spatial inhomogeneous periodic oscillation of prey and predator population densities.

| [1] |
E. F. D. Goufo, Y. Khan, Q. A. Chaudhry, HIV and shifting epicenters for COVID-19, an alert for some countries, Chaos Solitons Fract., 139 (2020), 110030. http://dx.doi.org/10.1016/j.chaos.2020.110030 doi: 10.1016/j.chaos.2020.110030

|
| [2] |
N. Faraz, Y. Khan, E. Goufo, A. Anjum, A. Anjum, Dynamic analysis of the mathematical model of COVID-19 with demographic effects, Z. Naturforsch C. J. Biosci., 75 (2020), 389–396. https://doi.org/10.1515/znc-2020-0121 doi: 10.1515/znc-2020-0121
|
| [3] |
Q. Xue, C. Liu, L. Li, G. Q. Sun, Z. Wang, Interactions of diffusion and nonlocal delay give rise to vegetation patterns in semi-arid environments, Appl. Math. Comput., 399 (2021), 126038. http://dx.doi.org/10.1016/j.amc.2021.126038 doi: 10.1016/j.amc.2021.126038
|
| [4] |
R. Yang, D. Jin, Dynamics in a predator-prey model with memory effect in predator and fear effect in prey, Electronic Res. Arch., 30 (2022), 1322–1339. http://dx.doi.org/10.3934/era.2022069 doi: 10.3934/era.2022069
|
| [5] |
R. Yang, C. Zhang, Dynamics in a diffusive predator-prey system with a constant prey refuge and delay, Nonlinear Anal.: Real World Appl., 31 (2016), 1–22. http://dx.doi.org/10.1016/j.nonrwa.2016.01.005 doi: 10.1016/j.nonrwa.2016.01.005
|
| [6] |
M. A. Khan, R. Khan, Y. Khan, S. Islam, A mathematical analysis of Pine Wilt disease with variable population size and optimal control strategies, Chaos Solitons Fract., 108 (2018), 205–217. http://dx.doi.org/10.1016/j.chaos.2018.02.002 doi: 10.1016/j.chaos.2018.02.002
|
| [7] |
Y. Khan, H. V$\acute{a}$zquez-Leal, Q. Wu, An efficient iterated method for mathematical biology model, Neural Comput. Applic., 23 (2013), 677–682. http://dx.doi.org/10.1007/s00521-012-0952-z doi: 10.1007/s00521-012-0952-z
|
| [8] |
Y. Khan, N. Faraz, Z. Smarda, An efficient iterated method for mathematical biology model, Neural Comput. Applic., 27 (2016), 671–675. http://dx.doi.org/10.1007/s00521-015-1886-z doi: 10.1007/s00521-015-1886-z
|
| [9] |
S. Mishra, R. K. Upadhyay, Exploring the cascading effect of fear on the foraging activities of prey in a three species Agroecosystem, Eur. Phys. J. Plus, 136 (2021), 974. http://dx.doi.org/10.1140/epjp/s13360-021-01936-5 doi: 10.1140/epjp/s13360-021-01936-5
|
| [10] |
R. Yang, Q. Song, Yong. An, Spatiotemporal dynamics in a predator-prey model with functional response increasing in both predator and prey densities, Mathematics, 10 (2022), 17. http://dx.doi.org/10.3390/math10010017 doi: 10.3390/math10010017
|
| [11] |
R. Yang, X. Zhao, Yong. An, Dynamical analysis of a delayed diffusive predator-prey model with additional food provided and anti-predator behavior, Mathematics, 10 (2022), 469. http://dx.doi.org/10.3390/math10030469 doi: 10.3390/math10030469
|
| [12] |
Y. Shao, Dynamics of an impulsive stochastic predator-prey system with the Beddington-DeAngelis functional response, Axioms, 10 (2021), 323. http://dx.doi.org/10.3390/axioms10040323 doi: 10.3390/axioms10040323
|
| [13] | R. Yang, F. Wang, D. Jin, Spatially inhomogeneous bifurcating periodic solutions induced by nonlocal competition in a predator-prey system with additional food, Math. Methods Appl. Sci., 2022. http://dx.doi.org/10.1002/mma.8349 |
| [14] |
R. Yang, D. Jin, W. Wang, A diffusive predator-prey model with generalist predator and time delay, AIMS Math., 7 (2022), 4574–4591. http://dx.doi.org/10.3934/math.2022255 doi: 10.3934/math.2022255
|
| [15] |
C. Qin, J. Du, Y. Hui, Dynamical behavior of a stochastic predator-prey model with Holling-type III functional response and infectious predator, AIMS Math., 7 (2022), 7403–7418. http://dx.doi.org/10.3934/math.2022413 doi: 10.3934/math.2022413
|
| [16] |
Y. Kang, L. Wedekin, Dynamics of a intraguild predation model with generalist or specialist predator, J. Math. Biol., 67 (2013), 1227–1259. http://dx.doi.org/10.1007/s00285-012-0584-z doi: 10.1007/s00285-012-0584-z
|
| [17] |
S. Madec, J. Casas, G. Barles, C. Suppo, Bistability induced by generalist natural enemies can reverse pest invasions, J. Math. Biol., 75 (2017), 543–575. http://dx.doi.org/10.1007/s00285-017-1093-x doi: 10.1007/s00285-017-1093-x
|
| [18] |
L. N. Guin, S. Acharya, Dynamic behaviour of a reaction-diffusion predator-prey model with both refuge and harvesting, Nonlinear Dyn., 88 (2017), 1501–1533. http://dx.doi.org/10.1007/s11071-016-3326-8 doi: 10.1007/s11071-016-3326-8
|
| [19] |
R. K. Upadhyay, R. Agrawal, Dynamics and responses of a predator-prey system with competitive interference and time delay, Nonlinear Dyn., 83 (2016), 821–837. http://dx.doi.org/10.1007/s11071-015-2370-0 doi: 10.1007/s11071-015-2370-0
|
| [20] |
F. Liu, R. Yang, L. Tang, Hopf bifurcation in a diffusive predator-prey model with competitive interference, Chaos Solitons Fract., 120 (2019), 250–258. http://dx.doi.org/10.1016/j.chaos.2019.01.029 doi: 10.1016/j.chaos.2019.01.029
|
| [21] |
N. F. Britton, Aggregation and the competitive exclusion principle, J. Theor. Biol., 136 (1989), 57–66. https://doi.org/10.1016/S0022-5193(89)80189-4 doi: 10.1016/S0022-5193(89)80189-4
|
| [22] |
J. Furter, M. Grinfeld, Local vs. non-local interactions in population dynamics, J. Math. Biol., 27 (1989), 65–80. http://dx.doi.org/10.1007/BF00276081 doi: 10.1007/BF00276081
|
| [23] |
S. Wu, Y. Song, Spatiotemporal dynamics of a diffusive predator-prey model with nonlocal effect and delay, Commun. Nonlinear Sci. Numer. Simul., 89 (2020), 105310. http://dx.doi.org/10.1016/j.cnsns.2020.105310 doi: 10.1016/j.cnsns.2020.105310
|
| [24] |
D. Geng, W. Jiang, Y. Lou, H. Wang, Spatiotemporal patterns in a diffusive predator-prey system with nonlocal intraspecific prey competition, Stud. Appl. Math., 2021 (2021), 396–432. http://dx.doi.org/10.1111/sapm.12444 doi: 10.1111/sapm.12444
|
| [25] |
S. Chen, J. Yu, Stability and bifurcation on predator-prey systems with nonlocal prey competition, Discrete Cont. Dyn. Syst., 38 (2018), 43–62. http://dx.doi.org/10.3934/dcds.2018002 doi: 10.3934/dcds.2018002
|
| [26] |
Y. Liu, D. Duan, B. Niu, Spatiotemporal dynamics in a diffusive predator-prey model with group defense and nonlocal competition, Appl. Math. Lett., 103 (2020), 106175. https://doi.org/10.1016/j.aml.2019.106175 doi: 10.1016/j.aml.2019.106175
|
| [27] | J. Wu, Theory and applications of partial functional differential equations, Springer, 1996. https://doi.org/10.1007/978-1-4612-4050-1 |
| [28] | B. D. Hassard, N. D. Kazarinoff, Y. H. Wan, Theory and applications of Hopf bifurcation, Cambridge University Press, 1981. |




Figures(5)

Chenxuan Nie, Dan Jin, Ruizhi Yang. Hopf bifurcation analysis in a delayed diffusive predator-prey system with nonlocal competition and generalist predator[J]. AIMS Mathematics, 2022, 7(7): 13344-13360. doi: 10.3934/math.2022737







 DownLoad:
DownLoad: