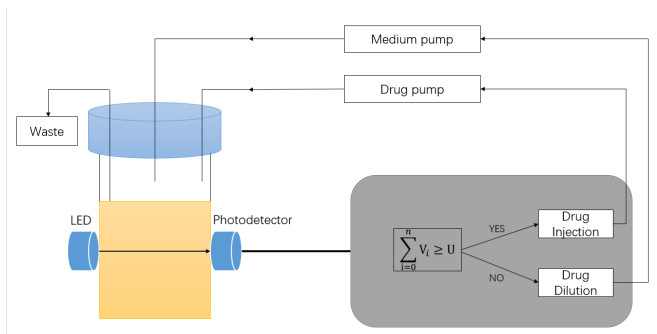
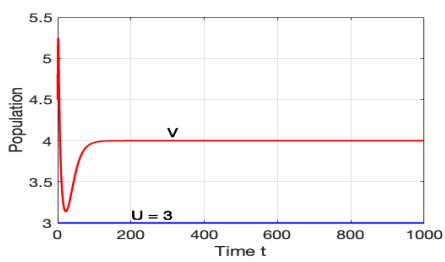
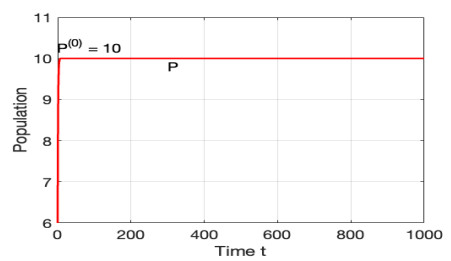
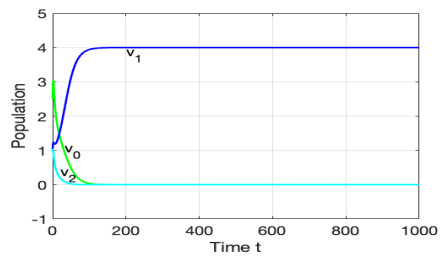
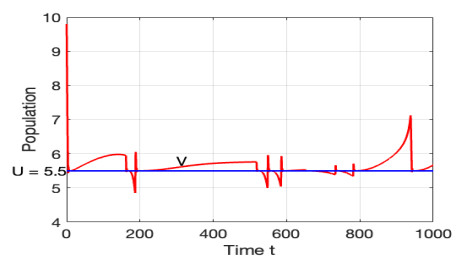
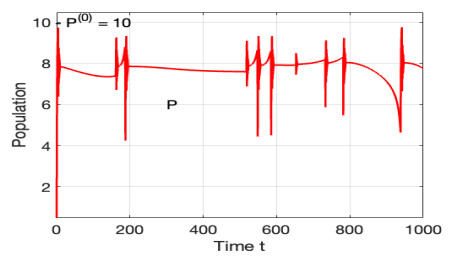
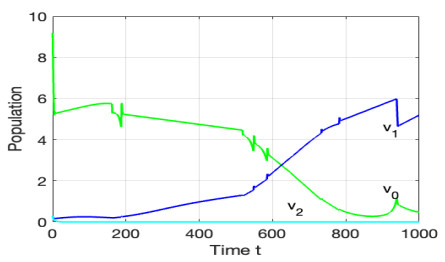
The morbidostat is a bacteria culture device that progressively increases antibiotic drug concentration. It is used to study the evolutionary pathway. In this article, we construct mathematical models for the morbidostat. First we consider the case of no mutations, we study limiting systems and obtain criteria for the large time behavior of the solutions. From the theoretical results and numerical simulations, we conclude that there are two competitive exclusion states of either wild type or mutant type as the threshold parameter $ U $ varies. There are three cases, wild type bacteria excludes all mutants; a mutant dominates in the competition; oscillation between the above two states.
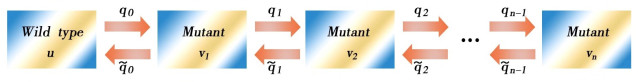
Next we study the systems of forward mutations and forward-backward mutations. Then we apply a result of pertubation for globally stable state.
Citation: Ziran Cheng, Sze-Bi Hsu. Dynamics of drug on-drug off models with mutations in morbidostat — Dedicated to the seventieth birthday of Professor Gail Wolkowicz[J]. AIMS Mathematics, 2023, 8(9): 20815-20840. doi: 10.3934/math.20231061
The morbidostat is a bacteria culture device that progressively increases antibiotic drug concentration. It is used to study the evolutionary pathway. In this article, we construct mathematical models for the morbidostat. First we consider the case of no mutations, we study limiting systems and obtain criteria for the large time behavior of the solutions. From the theoretical results and numerical simulations, we conclude that there are two competitive exclusion states of either wild type or mutant type as the threshold parameter $ U $ varies. There are three cases, wild type bacteria excludes all mutants; a mutant dominates in the competition; oscillation between the above two states.
Next we study the systems of forward mutations and forward-backward mutations. Then we apply a result of pertubation for globally stable state.

| [1] | H. L. Smith, P. E.Waltman, The theory of the chemostat, Cambridge University Press, Cambridge, 1995. https://doi.org/10.1017/CBO9780511530043 |
| [2] | S. B. Hsu, K. C. Chen, Ordinary differential equations with applications, Series on Applied Mathematics, Vol 23, World Scientific Press, 2022, 3rd Edition. https://doi.org/10.1142/8744 |
| [3] | W. A. Coppel, Stability and asymptotic behaivor of differential equations, Health. Math. Monograph, 1965. |
| [4] | P. Lancaster, M. Tismenetsky, The Theory of Matrices-second edition: With Applications, Academic Press, 1985. |
| [5] |
Z. Chen, S. B. Hsu, Y. T. Yang, The continuous Morbidostat : a chemostat controlled drug application to select for drug resistance mutants, Commun. Pur. Appl. Anal., 19 2020,203–220. https://doi.org/10.3934/cpaa.2020011 doi: 10.3934/cpaa.2020011

|
| [6] |
Z. Chen, S. B. Hsu, Y. T. Yang, The Morbidostat: A Bio-reactor That Promotes Selection for Drug Resistance in Bacteria, SIAM J. Appl. Math., 77 (2017), 470–499. https://doi.org/10.1137/16M105695X doi: 10.1137/16M105695X
|
| [7] |
H. L. Smith, P. Waltman, Perturbation of a Globally Stable Steady State, Proceedings of the American Mathematical Society, 127 (1999), 447–453. https://doi.org/10.1090/S0002-9939-99-04768-1 doi: 10.1090/S0002-9939-99-04768-1
|
| [8] |
M. Barber, Infection by penicillin resistant Staphylococci, Lancet, 2 (1948), 641–644. https://doi.org/10.1016/S0140-6736(48)92166-7 doi: 10.1016/S0140-6736(48)92166-7
|
| [9] |
S. B. Levy, B. Marshall, Antibacterial resistance worldwide: causes, challenges and responses, Nat. Med., 10 (2004), s122–s129. https://doi.org/10.1038/nm1145 doi: 10.1038/nm1145
|
| [10] |
E. Toprak, A. Veres, S. Yildiz, J. M. Pedraza, R. Chait, J. Paulsson, et al., Building a morbidostat: an automated continuous-culturre device for studying bacterial drug resistance under dynamically sustained drug inhibition, Nat. protoc., 8 (2013), 555–567. https://doi.org/10.1038/nprot.2013.021 doi: 10.1038/nprot.2013.021
|
| [11] | E. Toprak, A. Veres, J. B. Michel, R. Chait, D. L. Hartl, R. Kishony, Evolutionary paths to antibiotic resistance under dynamically sustained drug selection, Nat. Genet., 44 (2012), 101–105. |

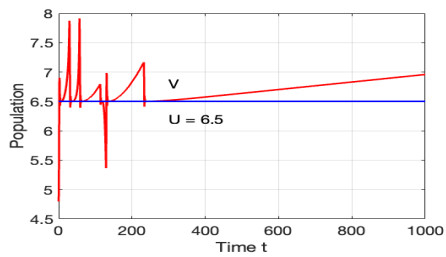
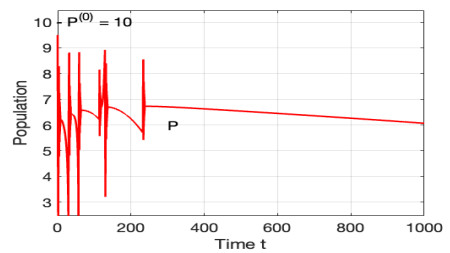
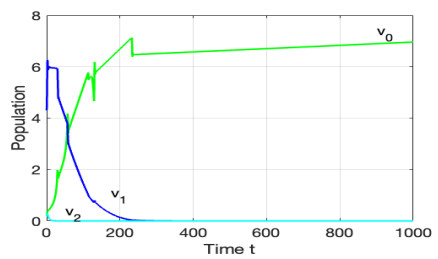
Figures(12)

Ziran Cheng, Sze-Bi Hsu. Dynamics of drug on-drug off models with mutations in morbidostat — Dedicated to the seventieth birthday of Professor Gail Wolkowicz[J]. AIMS Mathematics, 2023, 8(9): 20815-20840. doi: 10.3934/math.20231061







 DownLoad:
DownLoad: