Citation: A. M. Elaiw, N. H. AlShamrani, A. D. Hobiny. Mathematical modeling of HIV/HTLV co-infection with CTL-mediated immunity[J]. AIMS Mathematics, 2021, 6(2): 1634-1676. doi: 10.3934/math.2021098

| [1] | WHO, Global Health Observatory (GHO) data. HIV/AIDS, 2018. Available from: http://www.who.int/gho/hiv/en/. |
| [2] | M. A. Nowak, R. M. May, Virus dynamics: Mathematical principles of immunology and virology, Oxford University Press, 2001. |
| [3] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. doi: 10.1126/science.272.5258.74

|
| [4] |
K. D. Pedro, A. J. Henderson, L. M. Agosto, Mechanisms of HIV-1 cell-to-cell transmission and the establishment of the latent reservoir, Virus Res., 265 (2019), 115-121. doi: 10.1016/j.virusres.2019.03.014
|
| [5] | L. Rong, A. S. Perelson, Modeling latently infected cell activation: viral and latent reservoir persistence, and viral blips in HIV-infected patients on potent therapy, PLoS Comput. Biol., 5 (2009), 1-18. |
| [6] |
D. S. Callaway, A. S. Perelson, HIV-1 infection and low steady state viral loads, Bull. Math. Biol., 64 (2002), 29-64. doi: 10.1006/bulm.2001.0266
|
| [7] |
A. M. Elaiw, S. A. Azoz, Global properties of a class of HIV infection models with Beddington-DeAngelis functional response, Math. Method. Appl. Sci., 36 (2013), 383-394. doi: 10.1002/mma.2596
|
| [8] |
A. M. Elaiw, M. A. Alshaikh, Stability of discrete-time HIV dynamics models with three categories of infected CD4+ T-cells, Adv. Differ. Equ., 2019 (2019), 1-24. doi: 10.1186/s13662-018-1939-6
|
| [9] |
A. M. Elaiw, N. H. AlShamrani, Stability of a general CTL-mediated immunity HIV infection model with silent infected cell-to-cell spread, Adv. Differ. Equ., 2020 (2020), 1-25. doi: 10.1186/s13662-019-2438-0
|
| [10] |
A. M. Elaiw, E. K. Elnahary, A. A. Raezah, Effect of cellular reservoirs and delays on the global dynamics of HIV, Adv. Differ. Equ., 2018 (2018), 1-36. doi: 10.1186/s13662-017-1452-3
|
| [11] |
B. Buonomo, C. Vargas-De-Leon, Global stability for an HIV-1 infection model including an eclipse stage of infected cells, J. Math. Anal. Appl., 385 (2012), 709-720. doi: 10.1016/j.jmaa.2011.07.006
|
| [12] |
H. Liu, J. F. Zhang, Dynamics of two time delays differential equation model to HIV latent infection, Physica A, 514 (2019), 384-395. doi: 10.1016/j.physa.2018.09.087
|
| [13] | S. Iwami, J. S. Takeuchi, S. Nakaoka, F. Mammano, F. Clavel, H. Inaba, et al. Cell-to-cell infection by HIV contributes over half of virus infection, eLife, 4 (2015), 1-16. |
| [14] |
M. Sourisseau, N. Sol-Foulon, F. Porrot, F. Blanchet, O. Schwartz, Inefficient human immunodeficiency virus replication in mobile lymphocytes, J. Virol., 81 (2007), 1000-1012. doi: 10.1128/JVI.01629-06
|
| [15] |
A. Sigal, J. T. Kim, A. B. Balazs, E. Dekel, A. Mayo, R. Milo, et al. Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy, Nature, 477 (2011), 95-98. doi: 10.1038/nature10347
|
| [16] |
Y. Gao, J. Wang, Threshold dynamics of a delayed nonlocal reaction-diffusion HIV infection model with both cell-free and cell-to-cell transmissions, J. Math. Anal. Appl., 488 (2020), 124047. doi: 10.1016/j.jmaa.2020.124047
|
| [17] | A. Mojaver, H. Kheiri, Mathematical analysis of a class of HIV infection models of CD4+T-cells with combined antiretroviral therapy, Appl. Math. Comput., 259 (2015), 258-270. |
| [18] |
X. Wang, S. Tang, X. Song, L. Rong, Mathematical analysis of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission, J. Biol. Dynam., 11 (2017), 455-483. doi: 10.1080/17513758.2016.1242784
|
| [19] |
H. R. Norrgren, S. Bamba, O. Larsen, Z. Da Silva, P. Aaby, T. Koivula, et al. Increased prevalence of HTLV-1 in patients with pulmonary tuberculosis coinfected with HIV, but not in HIV-negative patients with tuberculosis, J. Acq. Imm. Def., 48 (2008), 607-610. doi: 10.1097/QAI.0b013e31817efb83
|
| [20] |
A. G. Lim, P. K. Maini, HTLV-Iinfection: A dynamic struggle between viral persistence and host immunity, J. Theor. Biol., 352 (2014), 92-108. doi: 10.1016/j.jtbi.2014.02.022
|
| [21] |
X. Pan, Y. Chen, H. Shu, Rich dynamics in a delayed HTLV-I infection model: Stability switch, multiple stable cycles, and torus, J. Math. Anal. Appl., 479 (2019), 2214-2235. doi: 10.1016/j.jmaa.2019.07.051
|
| [22] |
M. Y. Li, A. G. Lim, Modelling the role of Tax expression in HTLV-1 persisence in vivo, Bull. Math. Biol., 73 (2011), 3008-3029. doi: 10.1007/s11538-011-9657-1
|
| [23] |
B. Asquith, C. R. M. Bangham, The dynamics of T-cell fratricide: application of a robust approach to mathematical modeling in immunology, J. Theor. Biol., 222 (2003), 53-69. doi: 10.1016/S0022-5193(03)00013-4
|
| [24] | S. Tokudome et al., Incidence of adult T cell leukemia/lymphoma among human T lymphotropic virus type 1 carriers in Saga, Japan, Cancer Res., 49 (1989), 226-228. |
| [25] |
N. I. Stilianakis, J. Seydel, Modeling the T-cell dynamics and pathogenesis of HTLV-I infection, Bull. Math. Biol., 61 (1999), 935-947. doi: 10.1006/bulm.1999.0117
|
| [26] |
H. Gomez-Acevedo, M. Y. Li, Backward bifurcation in a model for HTLV-I infection of CD4+ T cells, Bull. Math. Biol., 67 (2005), 101-114. doi: 10.1016/j.bulm.2004.06.004
|
| [27] | C. Vargas-De-Leon, The complete classification for global dynamics of amodel for the persistence of HTLV-1 infection, Appl. Math. Comput., 237 (2014), 489-493. |
| [28] | X. Song, Y. Li, Global stability and periodic solution of a model for HTLV-1 infection and ATL progression, Appl. Math. Comput., 180 (2006), 401-410. |
| [29] |
E. S. Rosenberg, M. Altfeld, S. H. Poon, M. N. Phillips, B. M. Wilkes, R. L. Eldridge, et al., Immune control of HIV-1 following early treatment of acute infection, Nature, 407 (2000), 523-526. doi: 10.1038/35035103
|
| [30] |
A. M. Elaiw, N. H. AlShamrani, Global stability of a delayed adaptive immunity viral infection with two routes of infection and multi-stages of infected cells, Commun. Nonlinear Sci., 86 (2020), 105259. doi: 10.1016/j.cnsns.2020.105259
|
| [31] |
B. Asquith, C. R. M. Bangham, Quantifying HTLV-I dynamics, Immunol. Cell Biol., 85 (2007), 280-286. doi: 10.1038/sj.icb.7100050
|
| [32] |
C. Bartholdy, J. P. Christensen, D. Wodarz, A. R. Thomsen, Persistent virus infection despite chronic cytotoxic T-lymphocyte activation in gamma interferon-deficient mice infected with lymphocytic choriomeningitis virus, J. Virol., 74 (2000), 10304-10311. doi: 10.1128/JVI.74.22.10304-10311.2000
|
| [33] |
H. Gomez-Acevedo, M. Y. Li, S. Jacobson, Multi-stability in a model for CTL response to HTLV-I infection and its consequences Bull. Math. Biol., 72 (2010), 681-696. doi: 10.1007/s11538-009-9465-z
|
| [34] |
M. Y. Li, H. Shu, Multiple stable periodic oscillations in a mathematical model of CTL response to HTLV-I infection, Bull. Math. Biol., 73 (2011), 1774-1793. doi: 10.1007/s11538-010-9591-7
|
| [35] | L. Wang, Z. Liu, Y. Li, D. Xu, Complete dynamical analysis for a nonlinear HTLV-I infection model with distributed delay, CTL response and immune impairment, Discrete Cont. Dyn. B, 25 (2020), 917-933. |
| [36] |
Y. Wang, J. Liu, J. M. Heffernan, Viral dynamics of an HTLV-I infection model with intracellular delay and CTL immune response delay, J. Math. Anal. Appl., 459 (2018), 506-527. doi: 10.1016/j.jmaa.2017.10.027
|
| [37] |
F. Li, W. Ma, Dynamics analysis of an HTLV-1 infection model with mitotic division of actively infected cells and delayed CTL immune response, Math. Method. Appl. Sci., 41 (2018), 3000-3017. doi: 10.1002/mma.4797
|
| [38] |
S. Li, Y. Zhou, Backward bifurcation of an HTLV-I model with immune response, Discrete Cont. Dyn. B, 21 (2016), 863-881. doi: 10.3934/dcdsb.2016.21.863
|
| [39] | C. Casoli, E. Pilotti, U. Bertazzoni, Molecular and cellular interactions of HIV-1/HTLV coinfection and impact on AIDS progression, AIDS Rev., 9 (2007), 140-149. |
| [40] | E. Pilotti, M. V. Bianchi, A. De Maria, F. Bozzano, M. G. Romanelli, U. Bertazzoni, et al. HTLV-1/-2 and HIV-1 co-infections: retroviral interference on host immune status, Front. Microbiol., 4 (2013), 1-13. |
| [41] |
M. A. Beilke, K. P. Theall, M. O'Brien, J. L. Clayton, S. M. Benjamin, E. L. Winsor, et al. Clinical outcomes and disease progression among patients coinfected with HIV and human T lymphotropic virus types 1 and 2, Clin. Infect. Dis., 39 (2004), 256-263. doi: 10.1086/422146
|
| [42] |
C. Isache, M. Sands, N. Guzman, D. Figueroa, HTLV-1 and HIV-1 co-infection: A case report and review of the literature, IDCases, 4 (2016), 53-55. doi: 10.1016/j.idcr.2016.03.002
|
| [43] | M. Tulius Silva, O. de Melo Espíndola, A. C. Bezerra Leite, A. Araújo, Neurological aspects of HIV/human T lymphotropic virus coinfection, AIDS Rev., 11 (2009), 71-78. |
| [44] | N. Rockwood, L. Cook, H. Kagdi, S. Basnayake, C. R. M. Bangham, A. L. Pozniak, et al. Immune compromise in HIV-1/HTLV-1 coinfection with paradoxical resolution of CD4 lymphocytosis during antiretroviral therapy: A case report, Medicine, 94 (2015), 1-4. |
| [45] | C. Brites, J. Sampalo, A. Oliveira, HIV/human T-cell lymphotropic virus coinfection revisited: impact on AIDS progression, AIDS Rev., 11 (2009), 8-16. |
| [46] |
V. E. V. Geddes, D. P. José, F. E. Leal, D. F. Nixond, A. Tanuri, R. S. Aguiar, HTLV-1 Tax activates HIV-1 transcription in latency models, Virology, 504 (2017), 45-51. doi: 10.1016/j.virol.2017.01.014
|
| [47] |
R. Bingham, E. Dykeman, R. Twarock, RNA virus evolution via a quasispecies-based model reveals a drug target with a high barrier to resistance, Viruses, 9 (2017), 347. doi: 10.3390/v9110347
|
| [48] |
E. Ticona, M. A. Huaman, O. Yanque, J. R. Zunt, HIV and HTLV-1 coinfection: the need to initiate antiretroviral therapy, J. Int. Assoc. Provid. AIDS Care, 12 (2013), 373-374. doi: 10.1177/2325957413500988
|
| [49] | N. Bellomo, R. Bingham, M. A. J. Chaplain, G. Dosi, G. Forni, D. A. Knopoff, et al. A multi-scale model of virus pandemic: Heterogeneous interactive entities in a globally connected world, Math. Mod. Meth. Appl. S., 30 (2000), 1591-1651. |
| [50] |
A. Korobeinikov, Global properties of basic virus dynamics models, Bull. Math. Biol., 66 (2004), 879-883. doi: 10.1016/j.bulm.2004.02.001
|
| [51] | E. A. Barbashin, Introduction to the theory of stability, Wolters-Noordhoff, 1970. |
| [52] | J. P. LaSalle, The stability of dynamical systems, Philadelphia, SIAM, 1976. |
| [53] |
A. M. Lyapunov, The general problem of the stability of motion, Int. J. Control, 55 (1992), 531-534. doi: 10.1080/00207179208934253
|
| [54] | A. Vandormael, F. Rego, S. Danaviah, L. Carlos Junior Alcantara, D. R. Boulware, T. de Oliveira, CD4+ T-cell count may not be a useful strategy to monitor antiretroviral therapy response in HTLV-1/HIV co-infected patients, Curr. HIV Res., 15 (2017), 225-231. |
| [55] |
T. Inoue, T. Kajiwara, T. Saski, Global stability of models of humoral immunity against multiple viral strains, J. Biol. Dynam., 4 (2010), 282-295. doi: 10.1080/17513750903180275
|
| [56] |
G. Huang, Y. Takeuchi, W. Ma, Lyapunov functionals for delay differential equations model of viral infections, SIAM J. Appl. Math., 70 (2010), 2693-2708. doi: 10.1137/090780821
|
| [57] |
A. M. Elaiw, I. A. Hassanien, S. A. Azoz, Global stability of HIV infection models with intracellular delays, J. Korean Math. Soc., 49 (2012), 779-794. doi: 10.4134/JKMS.2012.49.4.779
|
| [58] |
A. M. Elaiw, S. F. Alshehaiween, Global stability of delay-distributed viral infection model with two modes of viral transmission and B-cell impairment, Math. Method. Appl. Sci., 43 (2020), 6677-6701. doi: 10.1002/mma.6408
|
| [59] |
P. W. Nelson, J. D. Murray, A. S. Perelson, A model of HIV-1 pathogenesis that includes an intracellular delay, Math. Biosci., 163 (2000), 201-215. doi: 10.1016/S0025-5564(99)00055-3
|
| [60] |
A. M. Elaiw, A. A. Raezah, Stability of general virus dynamics models with both cellular and viral infections and delays, Math. Method. Appl. Sci., 40 (2017), 5863-5880. doi: 10.1002/mma.4436
|
| [61] |
A. M. Elaiw, N. A. Almuallem, Global dynamics of delay-distributed HIV infection models with differential drug efficacy in cocirculating target cells, Math. Method. Appl. Sci., 39 (2016), 4-31. doi: 10.1002/mma.3453
|
| [62] | A. M. Elaiw, S. F. Alshehaiween, A. D. Hobiny, Global properties of a delay-distributed HIV dynamics model including impairment of B-cell functions, Mathematics, 7 (2019), 1-27. |
| [63] | A. M. Elaiw, E. K. Elnahary, Analysis of general humoral immunity HIV dynamics model with HAART and distributed delays, Mathematics, 7 (2019), 1-35. |
| [64] |
R. V. Culshaw, S. Ruan, G. Webb, A mathematical model of cell-to-cell spread of HIV-1 that includes a time delay, J. Math. Biol., 46 (2003), 425-444. doi: 10.1007/s00285-002-0191-5
|
| [65] |
N. Bellomo, K. J. Painter, Y. Tao, M. Winkler, Occurrence vs. Absence of taxis-driven instabilities in a May-Nowak model for virus infection, SIAM J. Appl. Math., 79 (2019), 1990-2010. doi: 10.1137/19M1250261
|
| [66] |
A. M. Elaiw, A. D. AlAgha, Global analysis of a reaction-diffusion within-host malaria infection model with adaptive immune response, Mathematics, 8 (2020), 1-32. doi: 10.3390/math8101793
|
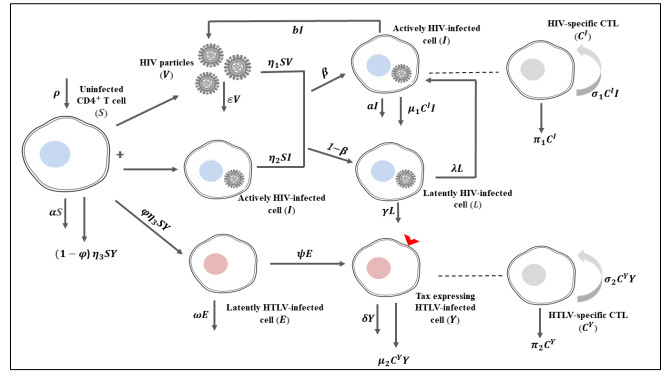
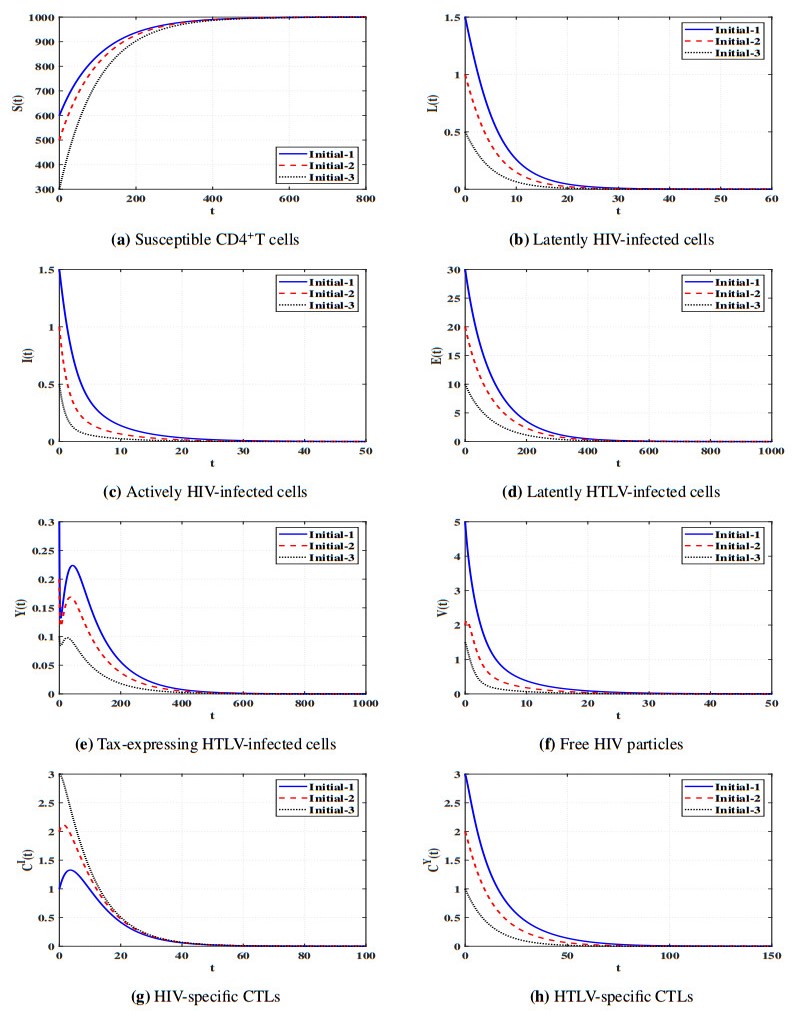
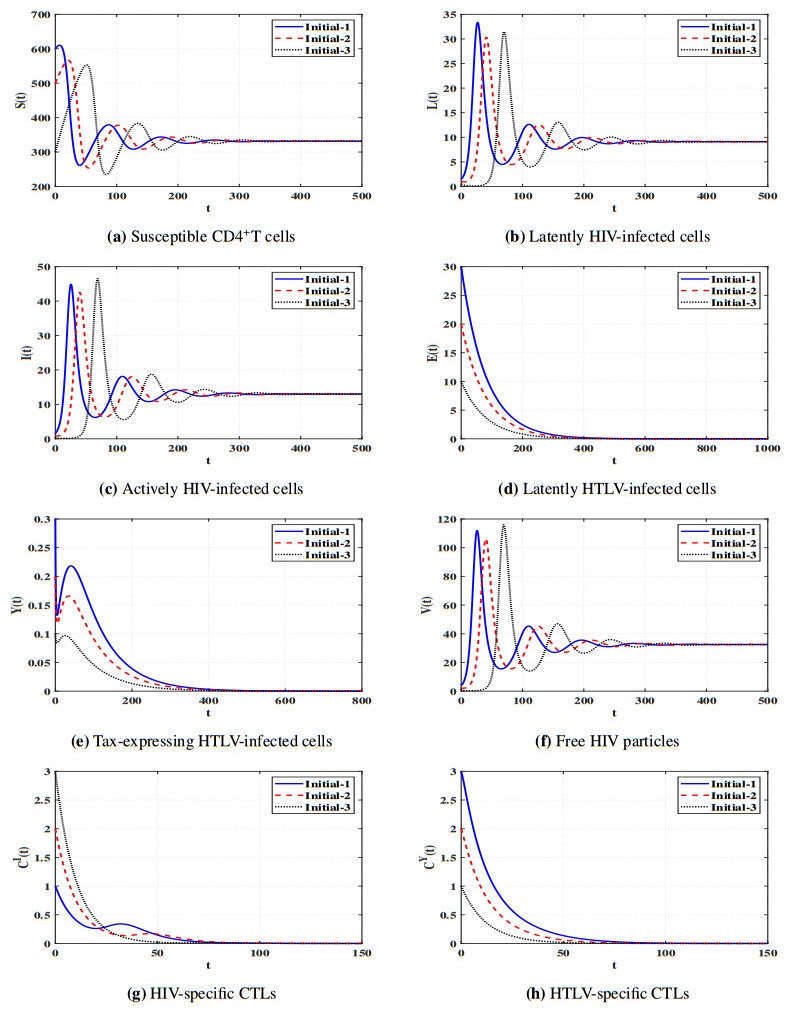
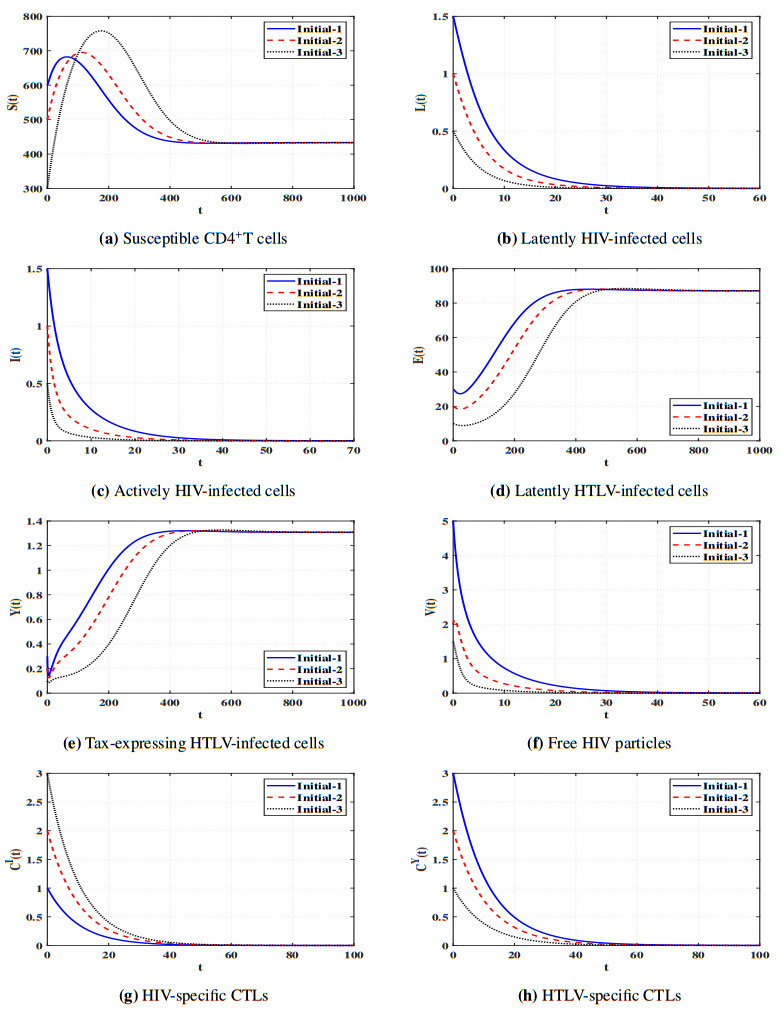
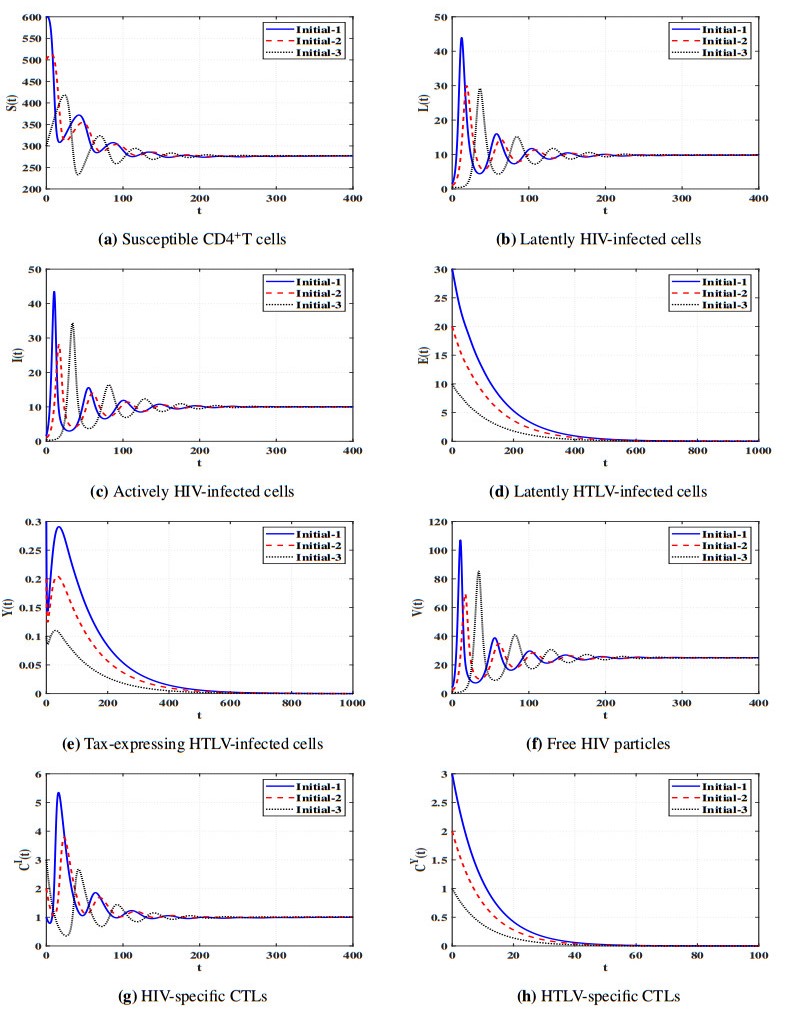
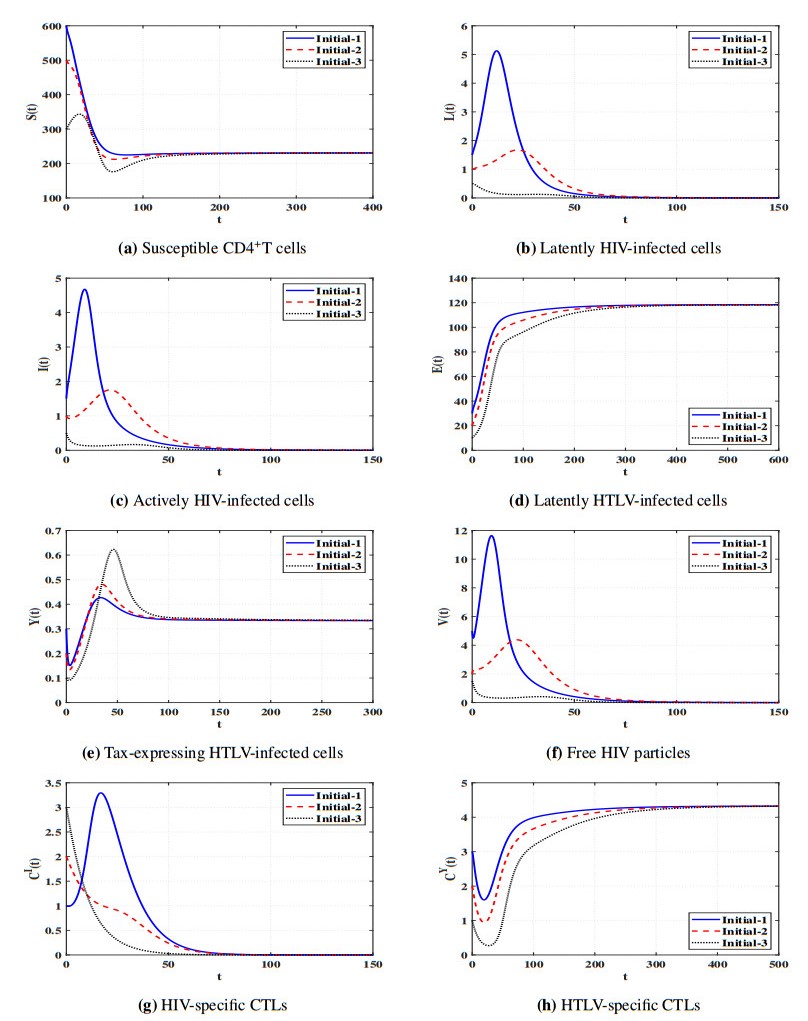
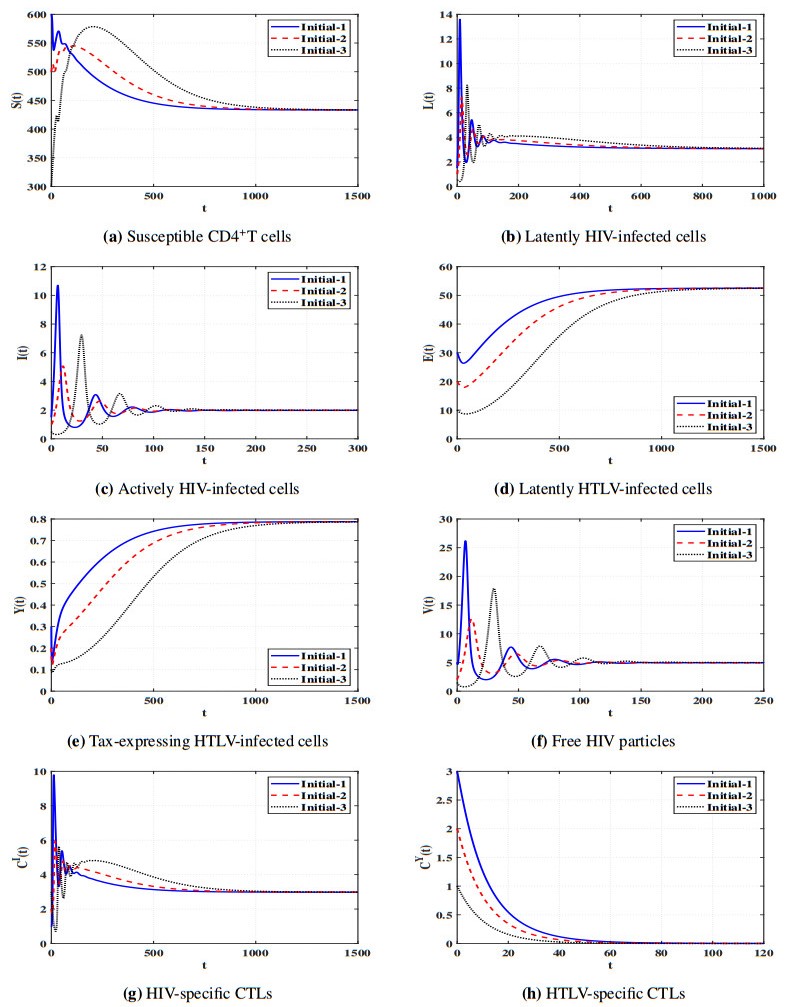
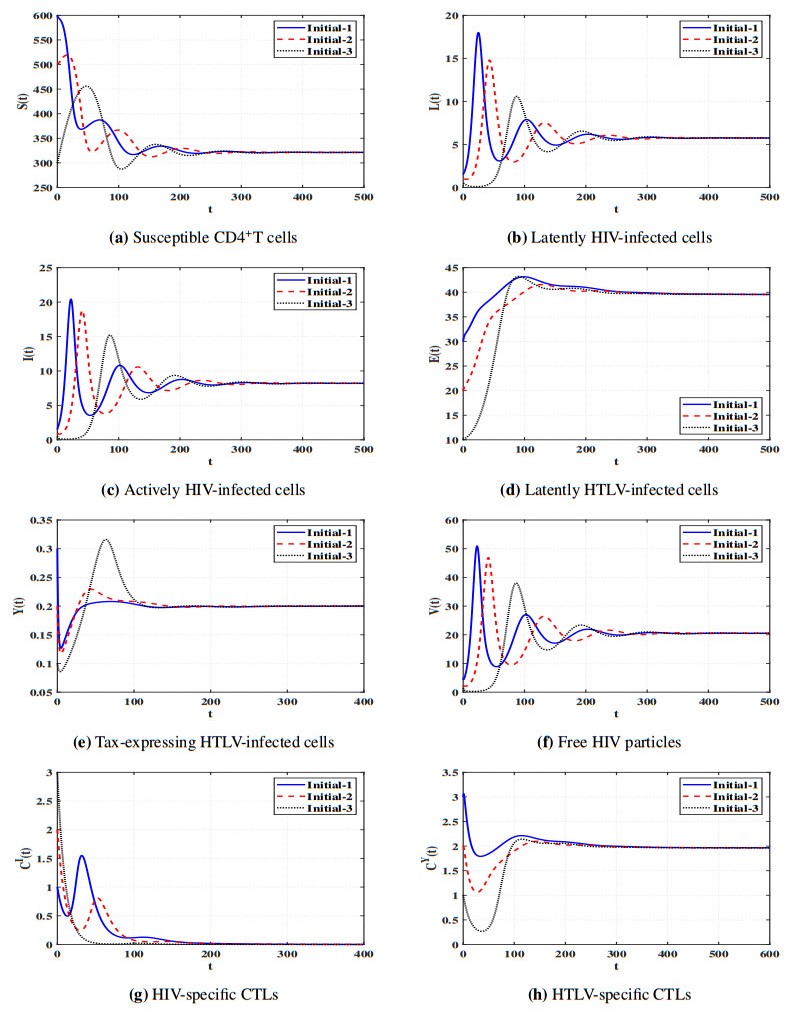
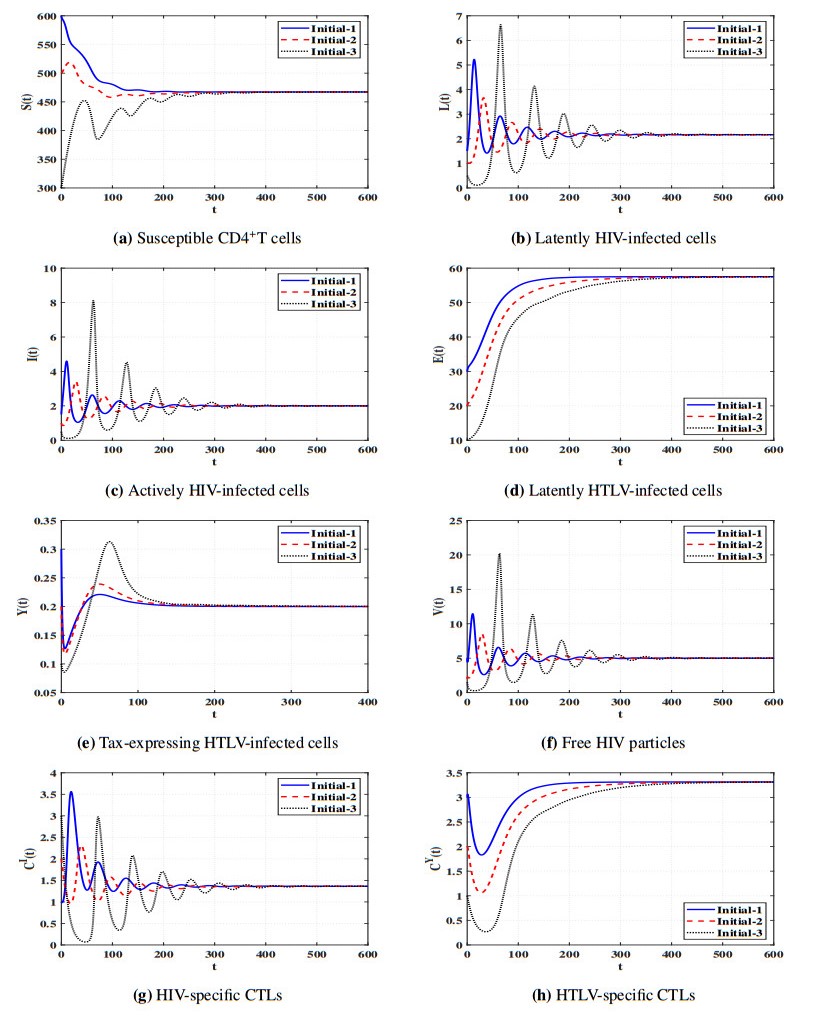
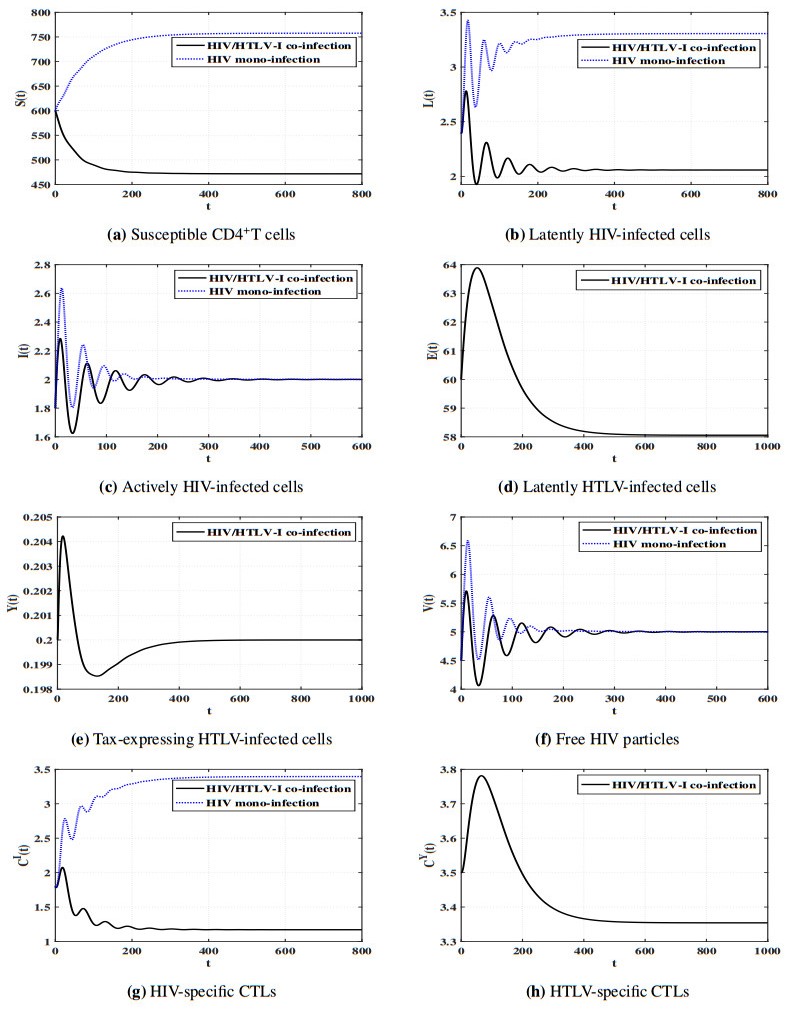
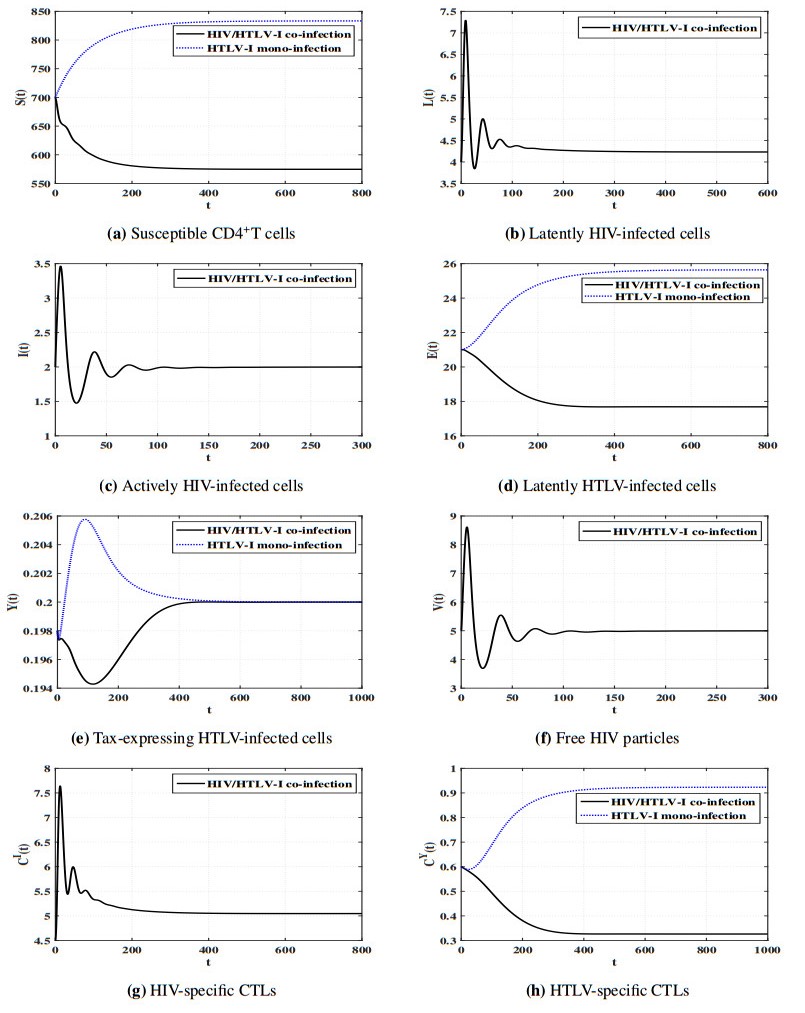
Figures(11) / Tables(5)

A. M. Elaiw, N. H. AlShamrani, A. D. Hobiny. Mathematical modeling of HIV/HTLV co-infection with CTL-mediated immunity[J]. AIMS Mathematics, 2021, 6(2): 1634-1676. doi: 10.3934/math.2021098







 DownLoad:
DownLoad: