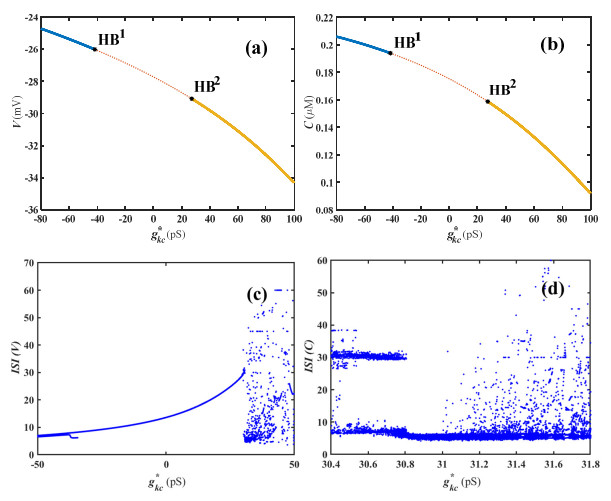
The central nervous system extensively expresses Ca2+-stimulated K+ channels, which serve to use Ca2+ to control their opening and closing. In this study, we explore the numerical computation of Hopf bifurcation in the Chay model based on the equilibrium point's stability and the center manifold theorem to illustrate the emergence of complicated neuronal bursting induced by variation of the conductance of the Ca2+-sensitive K+ channel. The results show that the formation and removal of various firing activities in this model are due to two subcritical Hopf bifurcations of equilibrium based on theoretical computation. Furthermore, the computational simulations are shown to support the validity of the conceptual approach. Consequently, the conclusion could be helpful to improve and deepen our understanding of the contribution of the Ca2+-sensitive K+ channel.
Citation: Danqi Feng, Yu Chen, Quanbao Ji. Contribution of a Ca2+-activated K+ channel to neuronal bursting activities in the Chay model[J]. Electronic Research Archive, 2023, 31(12): 7544-7555. doi: 10.3934/era.2023380
The central nervous system extensively expresses Ca2+-stimulated K+ channels, which serve to use Ca2+ to control their opening and closing. In this study, we explore the numerical computation of Hopf bifurcation in the Chay model based on the equilibrium point's stability and the center manifold theorem to illustrate the emergence of complicated neuronal bursting induced by variation of the conductance of the Ca2+-sensitive K+ channel. The results show that the formation and removal of various firing activities in this model are due to two subcritical Hopf bifurcations of equilibrium based on theoretical computation. Furthermore, the computational simulations are shown to support the validity of the conceptual approach. Consequently, the conclusion could be helpful to improve and deepen our understanding of the contribution of the Ca2+-sensitive K+ channel.

| [1] |
K. Koyama, H. Ando, K. Fujiwara, Functional improvement in β cell models of type 2 diabetes using on-demand feedback control, AIP Adv., 13 (2023), 045317. https://doi.org/10.1063/5.0124625 doi: 10.1063/5.0124625

|
| [2] |
Z. Yang, Q. Lu, L. Li, The genesis of period-adding bursting without bursting-chaos in the Chay model, Chaos, Solitons Fractals, 27 (2006), 689–697. https://doi.org/10.1016/j.chaos.2005.04.038 doi: 10.1016/j.chaos.2005.04.038
|
| [3] |
Z. Yang, Q. Lu, H. Gu, W. Ren, Integer multiple spiking in the stochastic Chay model and its dynamical generation mechanism, Phys. Lett. A, 299 (2002), 499–506. https://doi.org/10.1016/S0375-9601(02)00746-6 doi: 10.1016/S0375-9601(02)00746-6
|
| [4] |
M. Gu, Y. Zhu, X. Yin, D. M. Zhang, Small-conductance Ca2+-activated K+ channels: insights into their roles in cardiovascular disease, Exp. Mol. Med., 50 (2018), 1–7. https://doi.org/10.1038/s12276-018-0043-z doi: 10.1038/s12276-018-0043-z
|
| [5] |
X. Chen, Y. Feng, R. J. Quinn, D. L. Pountney, D. R. Richardson, G. D. Mellick, et al., Potassium channels in parkinson's disease: potential roles in its pathogenesis and innovative molecular targets for treatment, Pharm. Rev., 75 (2023), 758–788. https://doi.org/10.1124/pharmrev.122.000743 doi: 10.1124/pharmrev.122.000743
|
| [6] |
J. P. Adelman, J. Maylie, P. Sah, Small-conductance Ca2+-activated K+ channels: form and function, Annu. Rev. Physiol., 74 (2012), 245–269. https://doi.org/10.1146/annurev-physiol-020911-153336 doi: 10.1146/annurev-physiol-020911-153336
|
| [7] |
M. Al-Khannaq, J. Lytton, Regulation of K+-Dependent Na+/Ca2+-Exchangers (NCKX), Int. J. Mol. Sci., 24 (2023), 598. https://doi.org/10.3390/ijms24010598 doi: 10.3390/ijms24010598
|
| [8] |
S. Song, L. Luo, B. Sun, D. Sun, Roles of glial ion transporters in brain diseases, Glia, 68 (2020), 472–494. https://doi.org/10.1002/glia.23699 doi: 10.1002/glia.23699
|
| [9] |
Y. Li, R. Wang, T. Zhang, Nonlinear computational models of dynamical coding patterns in depression and normal rats: from electrophysiology to energy consumption, Nonlinear Dyn., 107 (2022), 3847–3862. https://doi.org/10.1007/s11071-021-07079-7 doi: 10.1007/s11071-021-07079-7
|
| [10] |
L. Li, Z. Zhao, White-noise-induced double coherence resonances in reduced Hodgkin-Huxley neuron model near subcritical Hopf bifurcation, Phys. Rev. E: Stat. Nonlinear Soft Matter Phys., 105 (2022), 034408. https://doi.org/10.1103/PhysRevE.105.034408 doi: 10.1103/PhysRevE.105.034408
|
| [11] |
P. Crotty, K. Segall, D. Schult, Biologically realistic behaviors from a superconducting neuron model, IEEE Trans. Appl. Supercond., 33 (2023), 1–6. https://doi.org/10.1109/TASC.2023.3242901 doi: 10.1109/TASC.2023.3242901
|
| [12] | L. Blomer, On the voltage gated ion channels involved in action potential generation and back propagation in layer 5 pyramidal neurons, 2022. Available from: https://theses.hal.science/tel-04077615/file/BLOMER_2022_archivage.pdf. |
| [13] |
M. V. Roshchin, V. N. Ierusalimsky, P. M. Balaban, E. S. Nikitin, Ca2+-activated KCa3.1 potassium channels contribute to the slow afterhyperpolarization in L5 neocortical pyramidal neurons, Sci. Rep., 10 (2020), 14484. https://doi.org/10.1038/s41598-020-71415-x doi: 10.1038/s41598-020-71415-x
|
| [14] |
R. Orfali, N. Albanyan, Ca2+-Sensitive potassium channels, Molecules, 28 (2023), 885. https://doi.org/10.3390/molecules28020885 doi: 10.3390/molecules28020885
|
| [15] |
L. Duan, Q. Lu, Q. Wang, Two-parameter bifurcation analysis of firing activities in the Chay neuronal model, Neurocomputing, 72 (2008), 341–351. https://doi.org/10.1016/j.neucom.2008.01.019 doi: 10.1016/j.neucom.2008.01.019
|
| [16] |
L. Duan, Q. Lu, Codimension-two bifurcation analysis on firing activities in Chay neuron model, Chaos, Solitons Fractals, 30 (2006), 1172–1179. https://doi.org/10.1016/j.chaos.2005.08.179 doi: 10.1016/j.chaos.2005.08.179
|
| [17] |
L. Duan, Q. Lu, Bursting oscillations near codimension-two bifurcations in the Chay Neuron model, Int. J. Nonlinear Sci. Numer. Simul., 7 (2006), 59–64. https://doi.org/10.1515/IJNSNS.2006.7.1.59 doi: 10.1515/IJNSNS.2006.7.1.59
|
| [18] |
Q. Xu, X. Tan, D. Zhu, H. Bao, Y. Hu, B. Bao, Bifurcations to bursting and spiking in the Chay neuron and their validation in a digital circuit, Chaos, Solitons Fractals, 141 (2020), 110353. https://doi.org/10.1016/j.chaos.2020.110353 doi: 10.1016/j.chaos.2020.110353
|
| [19] |
Q. Xu, X. Tan, D. Zhu, M. Chen, J. Zhou, H. Wu, Synchronous behavior for memristive synapse-connected Chay twin-neuron network and hardware implementation, Math. Probl. Eng., 2020 (2020), 8218740. https://doi.org/10.1155/2020/8218740 doi: 10.1155/2020/8218740
|
| [20] |
L. Lu, M. Yi, Z. Gao, Y. Wu, X. Zhao, Critical state of energy-efficient firing patterns with different bursting kinetics in temperature-sensitive Chay neuron, Nonlinear Dyn., 111 (2023), 16557–16567. https://doi.org/10.1007/s11071-023-08700-7 doi: 10.1007/s11071-023-08700-7
|
| [21] |
L. L. Lu, M. Yi, X. Q. Liu, Energy-efficient firing modes of chay neuron model in different bursting kinetics, Sci. China Technol. Sci., 65 (2022), 1661–1674. https://doi.org/10.1007/s11431-021-2066-7 doi: 10.1007/s11431-021-2066-7
|
| [22] |
F. Zhu, R. Wang, K. Aihara, X. Pan, Energy-efficient firing patterns with sparse bursts in the Chay neuron model, Nonlinear Dyn., 100 (2020), 2657–2672. https://doi.org/10.1007/s11071-020-05593-8 doi: 10.1007/s11071-020-05593-8
|
| [23] |
T. R. Chay, Chaos in a three-variable model of an excitable cell, Physica D, 16 (1985), 233–242. https://doi.org/10.1016/0167-2789(85)90060-0 doi: 10.1016/0167-2789(85)90060-0
|

Figures(2)
Danqi Feng, Yu Chen, Quanbao Ji. Contribution of a Ca2+-activated K+ channel to neuronal bursting activities in the Chay model[J]. Electronic Research Archive, 2023, 31(12): 7544-7555. doi: 10.3934/era.2023380







 DownLoad:
DownLoad: