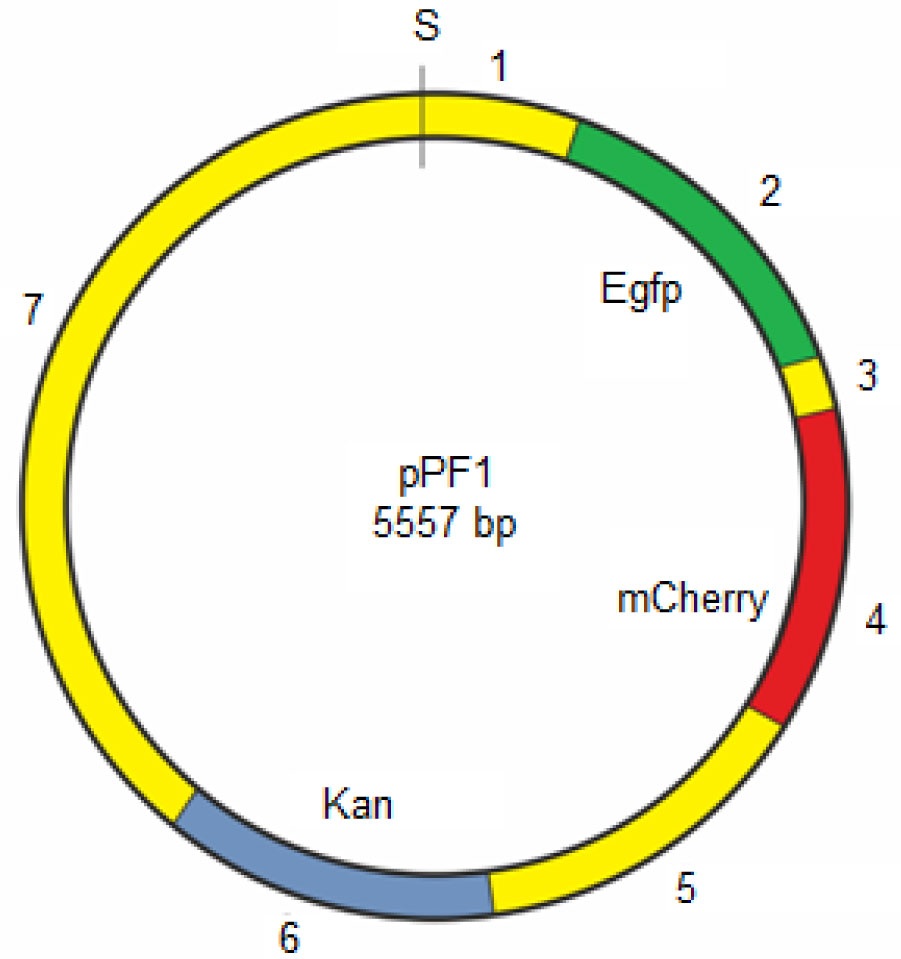
For better understanding the role of dynamic factors in the DNA functioning, it is important to study the internal mobility of DNA and, in particular, the movement of nonlinear conformational distortions -kinks along the DNA chains. In this work, we study the behavior of the kinks in the pPF1 plasmid containing two genes of fluorescent proteins (EGFP and mCherry). To simulate the movement, two coupled nonlinear sine-Gordon equations that describe the angular oscillations of nitrogenous bases in the main and complementary chains and take into account the effects of dissipation and the action of a constant torsion field. To solve the equations, approximate methods such as the quasi-homogeneous approximation, the mean field method, and the block method, were used. The obtained solutions indicate that two types of kinks moving along the double strand can be formed in any part of the plasmid. The profiles of the potential fields in which these kinks are moving are calculated. The results of the calculations show that the lowest energy required for the kink formation, corresponds to the region located between the genes of green and red proteins (EGFP and mCherry). It is shown that it is in this region a pit trap is located for both kinks. Trajectories of the kinks in the pit-trap and nearby are constructed. It is shown that there are threshold values of the torsion field, upon reaching which the kinks behavior changes dramatically: there is a transition from cyclic motion inside the pit-trap to translational motion and exit from the potential pit-trap.
Citation: Larisa A. Krasnobaeva, Ludmila V. Yakushevich. DNA kinks behavior in the potential pit-trap[J]. AIMS Biophysics, 2022, 9(2): 130-146. doi: 10.3934/biophy.2022012
For better understanding the role of dynamic factors in the DNA functioning, it is important to study the internal mobility of DNA and, in particular, the movement of nonlinear conformational distortions -kinks along the DNA chains. In this work, we study the behavior of the kinks in the pPF1 plasmid containing two genes of fluorescent proteins (EGFP and mCherry). To simulate the movement, two coupled nonlinear sine-Gordon equations that describe the angular oscillations of nitrogenous bases in the main and complementary chains and take into account the effects of dissipation and the action of a constant torsion field. To solve the equations, approximate methods such as the quasi-homogeneous approximation, the mean field method, and the block method, were used. The obtained solutions indicate that two types of kinks moving along the double strand can be formed in any part of the plasmid. The profiles of the potential fields in which these kinks are moving are calculated. The results of the calculations show that the lowest energy required for the kink formation, corresponds to the region located between the genes of green and red proteins (EGFP and mCherry). It is shown that it is in this region a pit trap is located for both kinks. Trajectories of the kinks in the pit-trap and nearby are constructed. It is shown that there are threshold values of the torsion field, upon reaching which the kinks behavior changes dramatically: there is a transition from cyclic motion inside the pit-trap to translational motion and exit from the potential pit-trap.

| [1] |
Zdravković S, Satarić MV, Daniel M (2013) Kink solitons in DNA. Int J Mod Phys B 27: 1350184. https://doi.org/10.1142/S0217979213501841 
|
| [2] |
Englander SW, Kallenbach NR, Heeger AJ, et al. (1980) Nature of the open state in long polynucleotide double helices: possibility of soliton excitations. P Natl Acad Sci USA 77: 7222-7226. https://doi.org/10.1073/pnas.77.12.7222
|
| [3] |
Hanke A, Metzler R (2003) Bubble dynamics in DNA. J Phys A: Math Gen 36: L473-L480. https://doi.org/10.1088/0305-4470/36/36/101
|
| [4] |
Altan-Bonnet G, Libchaber A, Krichevsky O (2003) Bubble Dynamics in double-stranded DNA. Phys Rev Lett 90: 138101-138105. https://doi.org/10.1103/PhysRevLett.90.138101
|
| [5] |
Okaly JB, Ndzana FII, Woulaché RL, et al. (2019) Base pairs opening and bubble transport in damped DNA dynamics with transport memory effects. Chaos: Interdiscipl J Nonlinear Sci 29: 093103. https://doi.org/10.1063/1.5098341
|
| [6] |
Shikhovtseva ES, Nazarov VN (2016) Non-linear longitudinal compression effect on dynamics of the transcription bubble in DAN. Biophys Chem 214–215: 47-53. https://doi.org/10.1016/j.bpc.2016.05.005
|
| [7] |
Grinevich AA, Ryasik AA, Yakushevich LV (2015) Trajectories of DNA bubbles. Chaos, Soliton Fract 75: 62-75. https://doi.org/10.1016/j.chaos.2015.02.009
|
| [8] |
Makasheva KA, Endutkin AV, Zharkov DO (2020) Requirements for DNA bubble structure for efficient cleavage by helix-two-turn-helix DNA glycosylases. Mutagenesis 35: 119-128. https://doi.org/10.1093/mutage/gez047
|
| [9] |
Hillebrand M, Kalosakas G, Bishop A R, et al. (2021) Bubble lifetimes in DNA gene promoters and their mutations affecting transcription. J Chem Phys 155: 095101. https://doi.org/10.1063/5.0060335
|
| [10] | The gfp green fluorescent protein [Neisseria gonorrhoeae] sequence, 2020. Available from: https://www.ncbi.nlm.nih.gov/gene/7011691 |
| [11] | The mCherry sequence and map. Available from: https://www.snapgene.com/resources/plasmid-files/?set=fluorescent_protein_genes_and_plasmids&plasmid=mCherry |
| [12] |
Masulis IS, Babaeva ZSh, Chernyshov SV, et al. (2015) Visualizing the activity of Escherichia coli divergent promoters and probing their dependence on superhelical density using dual-colour fluorescent reporter vector. Sci Rep 5: 11449. https://doi.org/10.1038/srep11449
|
| [13] | The pET-28b sequence and map. Available from: https://www.snapgene.com/resources/plasmid-files/?set=pet_and_duet_vectors_(novagen)&plasmid=pET-28b(%2B) |
| [14] |
Grinevich AA, Masulis IS, Yakushevich LV (2021) Mathematical modeling of transcription bubble behavior in the pPF1 plasmid and its modified versions: the link between the plasmid energy profile and the direction of transcription. Biophysics 66: 209-217.
|
| [15] |
McLaughlin DW, Scott AC (1978) Perturbation analysis of fluxon dynamics. Phys Rev A 18: 1652. https://doi.org/10.1103/PhysRevA.18.1652
|
| [16] | McLaughlin DW, Scott AC (1977) A multisoliton perturbation theory. Solitons in action . New York: Academic Press 201-256. |
| [17] |
Yakushevich LV, Krasnobaeva LA (2021) Ideas and methods of nonlinear mathematics and theoretical physics in DNA science: the McLaughlin-Scott equation and its application to study the DNA open state dynamics. Biophys Rev 13: 315-338. https://doi.org/10.1007/s12551-021-00801-0
|
| [18] |
Kornyshev AA, Wynveen A (2004) Nonlinear effects in the torsional adjustment of interacting DNA. Phys Rev E 69: 041905. https://doi.org/10.1103/PhysRevE.69.041905
|
| [19] |
Cherstvy AG, Kornyshev AA (2005) DNA melting in aggregates: impeded or facilitated?. J Phys Chem B 109: 13024-13029. https://doi.org/10.1021/jp051117i
|
| [20] |
Peyrard M (2004) Nonlinear dynamics and statistical physics of DNA. Nonlinearity 17: R1. https://doi.org/10.1088/0951-7715/17/2/R01
|
| [21] |
Yakushevich LV, Krasnobaeva LA (2021) Double energy profile of pBR322 plasmid. AIMS Biophys 8: 221-232. https://doi.org/10.3934/biophy.2021016
|
 biophy-09-02-012-s001.pdf biophy-09-02-012-s001.pdf |
 |








Figures(9) / Tables(4)

Larisa A. Krasnobaeva, Ludmila V. Yakushevich. DNA kinks behavior in the potential pit-trap[J]. AIMS Biophysics, 2022, 9(2): 130-146. doi: 10.3934/biophy.2022012







 DownLoad:
DownLoad: