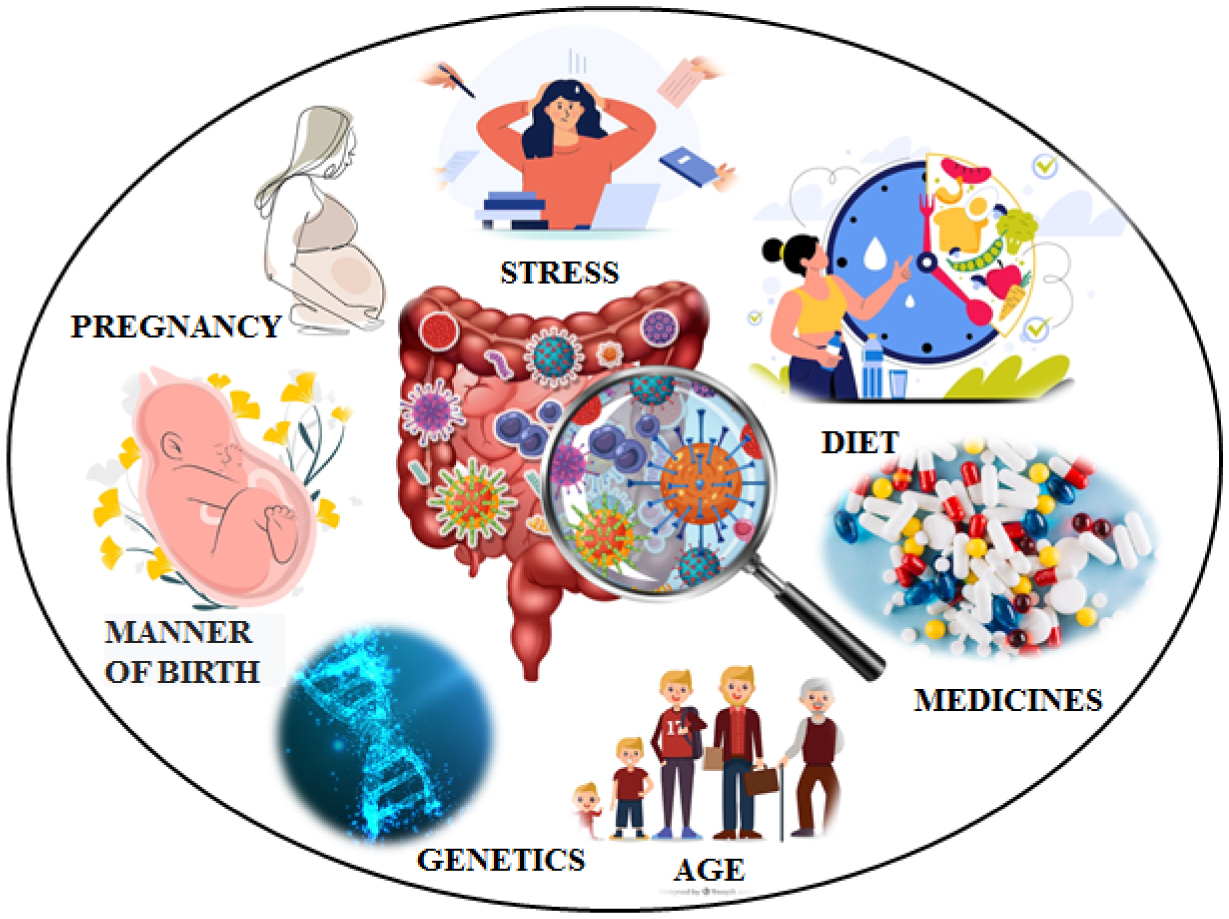
The study of the brain-gut axis and its impact on cognitive function and in the development of neurodegenerative diseases is a very timely topic of interest to researchers. This review summarizes information on the basic mechanisms of gut-brain communication. We then discuss the roles of the gut microbiome as a neuroprotective factor in neurodegeneration. The gut microbiota is extremely important in maintaining the body's homeostasis, shaping the human immune system and the proper functioning of the brain. The intestinal microflora affects the processes of neuroplasticity, synaptogenesis, and neuronal regeneration. This review aims to explain changes in the composition of the bacterial population of the intestinal microflora among patients with Alzheimer's disease, Parkinson's disease, and multiple sclerosis. Abnormalities in gut microflora composition are also noted in stress, depression, or autism spectrum development. New observations on psychobiotic supplementation in alleviating the symptoms of neurodegenerative diseases are also presented.
Citation: mgr Natalia Białoń, dr hab. n. o zdr. Dariusz Górka, mgr Mikołaj Górka. The brain-gut axis: communication mechanisms and the role of the microbiome as a neuroprotective factor in the development of neurodegenerative diseases: A literature overview[J]. AIMS Neuroscience, 2024, 11(3): 289-311. doi: 10.3934/Neuroscience.2024019
The study of the brain-gut axis and its impact on cognitive function and in the development of neurodegenerative diseases is a very timely topic of interest to researchers. This review summarizes information on the basic mechanisms of gut-brain communication. We then discuss the roles of the gut microbiome as a neuroprotective factor in neurodegeneration. The gut microbiota is extremely important in maintaining the body's homeostasis, shaping the human immune system and the proper functioning of the brain. The intestinal microflora affects the processes of neuroplasticity, synaptogenesis, and neuronal regeneration. This review aims to explain changes in the composition of the bacterial population of the intestinal microflora among patients with Alzheimer's disease, Parkinson's disease, and multiple sclerosis. Abnormalities in gut microflora composition are also noted in stress, depression, or autism spectrum development. New observations on psychobiotic supplementation in alleviating the symptoms of neurodegenerative diseases are also presented.

| [1] | Żakowicz J, Bramorska A, Zarzycka W, et al. (2020) Wpływ mikrobioty jelitowej na mózg, funkcje poznawcze i emocje. Kosmos problemy nauk biologicznych 69: 45-58. https://doi.org/10.36921/kos.2020_2634 |
| [2] |
Dinan TG, Stanton C, Cryan JF (2013) Psychobiotics: a novel class of psychotropic. Biol Psychiatry 74: 720-726. https://doi.org/10.1016/j.biopsych.2013.05.001 
|
| [3] |
Góralczyk-Bińkowska A, Szmajda-Krygier D, Kozłowska E (2022) The Microbiota-Gut-Brain Axis in Psychiatric Disorders. Int J Mol Sci 23: 11245. https://doi.org/10.3390/ijms231911245
|
| [4] |
Dinan TG, Cryan JF (2017) Brain-Gut-Microbiota Axis and Mental Health. Psychosom Med 79: 920-926. https://doi.org/10.1097/PSY.0000000000000519
|
| [5] |
Nowicka D, Chilicka K, Dzieńdziora-Urbińska I (2022) Host-Microbe Interaction on the Skin and Its Role in the Pathogenesis and Treatment of Atopic Dermatitis. Pathogens 11: 71. https://doi.org/10.3390/pathogens11010071
|
| [6] |
Daniel H (2020) Diet and the gut microbiome: From hype to hypothesis. Br J Nutr 124: 521-530. https://doi.org/10.1017/S0007114520001142
|
| [7] |
Enaud R, Vandenborght LE, Coron N, et al. (2018) The Mycobiome: A Neglected Component in the Microbiota-Gut-Brain Axis. Microorganisms 6: 22. https://doi.org/10.3390/microorganisms6010022
|
| [8] |
Sureda A, Daglia M, Argüelles Castilla S, et al. (2020) Oral microbiota and Alzheimer's disease: Do all roads lead to Rome?. Pharmacol Res 151: 104582. https://doi.org/10.1016/j.phrs.2019.104582
|
| [9] | Gałecka M, Basińska A, Bartnicka A (2018) Znaczenie mikrobioty jelitowej w kształtowaniu zdrowia człowieka – implikacje w praktyce lekarza rodzinnego. Journals Via Medica 12: 50-59. |
| [10] | Bartnicka A, Gałęcka M, Mazela J (2016) Wpływ czynników prenatalnych i postnatalnych na mikrobiotę jelitową noworodków. Stand Med Pediatr 13: 112-116. |
| [11] |
Dominguez-Bello MG, Costello EK, Contreras M, et al. (2010) Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A 107: 11971-11975. https://doi.org/10.1073/pnas.1002601107
|
| [12] | Cukrowska B (2016) Znaczenie programowania mikrobiotycznego w rozwoju przewlekłych chorób infekcyjnych. Stand Med Pediatr 13: 1019-1028. |
| [13] | Gałęcka M, Bartnicka A, Szewc M, et al. (2016) Kształtowanie się mikrobioty jelitowej u niemowląt warunkiem zachowania zdrowia. Standardy medyczne pediatria 13: 359-367. |
| [14] |
Lee YK, Menezes JS, Umesaki Y, et al. (2011) Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc Natl Acad Sci U S A : 4615-4622. https://doi.org/10.1073/pnas.1000082107
|
| [15] |
Tlaskalová-Hogenová H, Stěpánková R, Kozáková H, et al. (2011) The role of gut microbiota (commensal bacteria) and the mucosal barrier in the pathogenesis of inflammatory and autoimmune diseases and cancer: contribution of germ-free and gnotobiotic animal models of human diseases. Cell Mol Immunol 8: 110-120. https://doi.org/10.1038/cmi.2010.67
|
| [16] |
Martin CR, Osadchiy V, Kalani A, et al. (2018) The brain-gut-microbiome axis. Cell Mol Gastroenterol Hepatol 6: 133-148. https://doi.org/10.1016/j.jcmgh.2018.04.003
|
| [17] |
Brown K, DeCoffe D, Molcan E, et al. (2012) Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 4: 1095-1119. https://doi.org/10.3390/nu4081095
|
| [18] |
González Olmo BM, Butler MJ, Barrientos RM (2021) Evolution of the Human Diet and Its Impact on Gut Microbiota, Immune Responses, and Brain Health. Nutrients 13: 196. https://doi.org/10.3390/nu13010196
|
| [19] |
Foster JA, Rinaman L, Cryan JF (2017) Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol Stress 7: 124-136. https://doi.org/10.1016/j.ynstr.2017.03.001
|
| [20] |
Tannock GW, Savage DC (1974) Influences of dietary and environmental stress on microbial populations in the murine gastrointestinal tract. Infect Immun 9: 591-598. https://doi.org/10.1128/iai.9.3.591-598.1974
|
| [21] |
Bailey MT, Dowd SE, Galley JD, et al. (2011) Exposure to social stressors alters the structure of the intestinal microbiota. Brain Behav Immun 25: 397-407. https://doi.org/10.1016/j.bbi.2010.10.023
|
| [22] |
Cryan JF, O'Riordan KJ, Cowan CSM, et al. (2019) The Microbiota-Gut-Brain Axis. Physiol Rev 99: 1877-2013. https://doi.org/10.1152/physrev.00018.2018
|
| [23] |
Berthoud HR, Neuhuber WL (2000) Functional and chemical anatomy of the afferent vagal system. Auton Neurosci 85: 1-17. https://doi.org/10.1016/S1566-0702(00)00215-0
|
| [24] |
AGOSTONI E, CHINNOCK JE, DE DALY MB, et al. (1957) Functional and histological studies of the vagus nerve and its branches to the heart, lungs and abdominal viscera in the cat. J Physiol 135: 182-205. https://doi.org/10.1113/jphysiol.1957.sp005703
|
| [25] |
Prechtl JC, Powley TL (1990) B-Afferents: A fundamental division of the nervous system mediating homeostasis?. Behav Brain Sci 13: 289-300. https://doi.org/10.1017/S0140525X00078729
|
| [26] |
Suarez AN, Noble EE, Kanoski SE (2019) Regulation of memory function by feeding-relevant biological systems: Following the bread-crumbs to the hippocampus. Front Mol Neurosci 12: 1-21. https://doi.org/10.3389/fnmol.2019.00101
|
| [27] |
Wang Y, Zhan G, Cai Z, et al. (2021) Vagus nerve stimulation in brain diseases: Therapeutic applications and biological mechanisms. Neurosci Biobehav Rev 127: 37-53. https://doi.org/10.1016/j.neubiorev.2021.04.018
|
| [28] |
Jin Z, Dong J, Wang Y, et al. (2023) Exploring the potential of vagus nerve stimulation in treating brain diseases: a review of immunologic benefits and neuroprotective efficacy. Eur J Med Res 28: 444. https://doi.org/10.1186/s40001-023-01439-2
|
| [29] |
Bonaz B, Sinniger V, Pellissier S (2016) Anti-inflammatory properties of the vagus nerve: potential therapeutic implications of vagus nerve stimulation. J Physiol 594: 5781-5790. https://doi.org/10.1113/JP271539
|
| [30] |
Meregnani J, Clarençon D, Vivier M, et al. (2011) Anti-inflammatory effect of vagus nerve stimulation in a rat model of inflammatory bowel disease. Auton Neurosci 160: 82-89. https://doi.org/10.1016/j.autneu.2010.10.007
|
| [31] |
Merrill CA, Jonsson MA, Minthon L, et al. (2006) Vagus nerve stimulation in patients with Alzheimer's disease: additional follow-up results of a pilot study through 1 year. J Clin Psychiatry 67: 1171-1178. https://doi.org/10.4088/JCP.v67n0801
|
| [32] |
Dawson J, Engineer ND, Prudente CN, et al. (2020) Vagus nerve stimulation paired with upper-limb rehabilitation after stroke: one-year follow-up. Neurorehabil Neural Repair 34: 609-615. https://doi.org/10.1177/1545968320924361
|
| [33] |
Mondal B, Choudhury S, Simon B, et al. (2019) Noninvasive vagus nerve stimulation improves gait and reduces freezing of gait in Parkinson's disease. Mov Disord 34: 917-918. https://doi.org/10.1002/mds.27662
|
| [34] |
Hays SA (2016) Enhancing rehabilitative therapies with vagus nerve stimulation. Neurotherapeutics 13: 382-394. https://doi.org/10.1007/s13311-015-0417-z
|
| [35] |
Kaelberer MM, Buchanan K, Klein ME, et al. (2018) A gut-brain neural circuit for nutrient sensory transduction. Science 361. https://doi.org/10.1126/science.aat5236
|
| [36] |
Dinan TG, Stilling RM, Stanton C, et al. (2015) Collective unconscious: How gut microbes shape human behavior. J Psychiatric Res 63: 1-9. https://doi.org/10.1016/j.jpsychires.2015.02.021
|
| [37] |
Dantzer R (2009) Cytokine, sickness behavior, and depression. Immunol. Allergy Clin North Am 29: 247-264. https://doi.org/10.1016/j.iac.2009.02.002
|
| [38] |
Rogers GB, Keating DJ, Young RL, et al. (2016) From gut dysbiosis to altered brain function and mental illness: mechanisms and pathways. Mol Psychiatry 21: 738-748. https://doi.org/10.1038/mp.2016.50
|
| [39] |
Sarkar A, Lehto SM, Harty S, et al. (2016) Psychobiotics and the manipulation of bacteria-gut-brain signals. Trends Neurosci 39: 763-781. https://doi.org/10.1016/j.tins.2016.09.002
|
| [40] |
Martin-Gallausiaux C, Marinelli L, Blottière HM, et al. (2020) SCFA: mechanisms and functional importance in the gut. P Nutr Soc 80: 37-49. https://doi.org/10.1017/S0029665120006916
|
| [41] |
Fusco W, Lorenzo MB, Cintoni M, et al. (2011) Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 15: 2211. https://doi.org/10.3390/nu15092211
|
| [42] |
Atarashi K, Tanoue T, Shima T, et al. (2011) Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331: 337-341. https://doi.org/10.1126/science.1198469
|
| [43] |
Smith PM, Howitt MR, Panikov N, et al. (2013) The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341: 569-573. https://doi.org/10.1126/science.1241165
|
| [44] |
Dupraz L, Magniez A, Rolhion N, et al. (2021) Gut microbiota-derived short-chain fatty acids regulate IL-17 production by mouse and human intestinal γδ T cells. Cell Rep 36: 109332. https://doi.org/10.1016/j.celrep.2021.109332
|
| [45] |
Biagioli M, Carino A (2019) Signaling from intestine to the host: how bile acids regulate intestinal and liver immunity. Handb Exp Pharmacol 256: 95-108. https://doi.org/10.1007/164_2019_225
|
| [46] |
Bairamian D, Sha S, Rolhion N, et al. (2022) Microbiota in neuroinflammation and synaptic dysfunction: a focus on Alzheimer's disease. Mol Neurodegener 17: 19. https://doi.org/10.1186/s13024-022-00522-2
|
| [47] |
Hoogland ICM, Houbolt C, Westerloo DJ, et al. (2015) Systemic inflammation and microglial activation: systemic review of animal experiments. J Neuroinflamm 12: 114. https://doi.org/10.1186/s12974-015-0332-6
|
| [48] |
Gareau MG, Wine E, Rodrigues DM, et al. (2011) Bacterial infection causes stress-induced memory dysfunction in mice. Gut 60: 307-317. https://doi.org/10.1136/gut.2009.202515
|
| [49] |
Sudo N, Chida Y, Aiba Y, et al. (2004) Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol 558: 263-275. https://doi.org/10.1113/jphysiol.2004.063388
|
| [50] |
Silva YP, Bernardi A, Frozza RL (2020) The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol (Lausanne) 11: 25. https://doi.org/10.3389/fendo.2020.00025
|
| [51] |
Mao YK, Kasper DL, Wang B, et al. (2013) Bacteroides fragilis polysaccharide A is necessary and sufficient for acute activation of intestinal sensory neurons. Nat Comm 4: 1465. https://doi.org/10.1038/ncomms2478
|
| [52] |
De Vadder F, Grasset E, Holm LM, et al. (2018) Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc Natl Acad Sci USA 115: 6458-6463. https://doi.org/10.1073/pnas.1720017115
|
| [53] |
Nandwana V, Nandwana NK, Das Y, et al. (2022) The Role of Microbiome in Brain Development and Neurodegenerative Diseases. Molecules 27: 3402. https://doi.org/10.3390/molecules27113402
|
| [54] |
Wojtania MA, Majewska EM, Duda W (2024) Importance of gut microbiome in Alzheimer's disease. Med Srod 27: 60-65. https://doi.org/10.26444/ms/188315
|
| [55] |
Uysal G, Ozturk M (2020) Hippocampal atrophy based Alzheimer's disease diagnosis via machine learning methods. J Neurosci Methods 337: 108669. https://doi.org/10.1016/j.jneumeth.2020.108669
|
| [56] | Alzheimer's Association.Alzheimer's disease facts and figures. Alzheimers Dement (2016) 12: 459-509. https://doi.org/10.1016/j.jalz.2016.03.001 |
| [57] |
Winblad B, Amouyel P, Andrieu S, et al. (2016) Defeating Alzheimer's disease and other dementias: a priority for European science and society. Lancet Neurol 15: 455-532. https://doi.org/10.1016/S1474-4422(16)00062-4
|
| [58] |
Liu P, Wu L, Peng G, et al. (2019) Altered microbiomes distinguish Alzheimer's disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain Behav Immun 80: 633-643. https://doi.org/10.1016/j.bbi.2019.05.008
|
| [59] |
Roubaud-Baudron C, Krolak-Salmon P, Quadrio I, et al. (2012) Impact of chronic Helicobacter pylori infection on Alzheimer's disease: preliminary results. Neurobiol Aging 33: 1009.e11-9. https://doi.org/10.1016/j.neurobiolaging.2011.10.021
|
| [60] |
Chen SG, Stribinskis V, Rane MJ, et al. (2016) Exposure to the Functional Bacterial Amyloid Protein Curli Enhances Alpha-Synuclein Aggregation in Aged Fischer 344 Rats and Caenorhabditis elegans. Sci Rep 6: 34477. https://doi.org/10.1038/srep34477
|
| [61] |
Pistollato F, Sumalla Cano S, Elio I, et al. (2016) Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr Rev 74: 624-34. https://doi.org/10.1093/nutrit/nuw023
|
| [62] |
Jang SE, Lim SM, Jeong JJ, et al. (2018) Gastrointestinal inflammation by gut microbiota disturbance induces memory impairment in mice. Mucosal Immunol 11: 369-379. https://doi.org/10.1038/mi.2017.49
|
| [63] |
Zhao Y, Jaber V, Lukiw WJ (2017) Secretory Products of the Human GI Tract Microbiome and Their Potential Impact on Alzheimer's Disease (AD): Detection of Lipopolysaccharide (LPS) in AD Hippocampus. Front Cell Infect Microbiol 7: 318. https://doi.org/10.3389/fcimb.2017.00318
|
| [64] |
Sun MF, Shen YQ (2018) Dysbiosis of gut microbiota and microbial metabolites in Parkinson's disease. Ageing Res Rev 45: 53-61. https://doi.org/10.1016/j.arr.2018.04.004
|
| [65] |
Li B, He Y, Ma J, et al. (2019) Mild cognitive impairment has similar alterations as Alzheimer's disease in gut microbiota. Alzheimers Dement 15: 1357-1366. https://doi.org/10.1016/j.jalz.2019.07.002
|
| [66] |
Perez-Pardo P, Kliest T, Dodiya HB, et al. (2017) The gut-brain axis in Parkinson's disease: Possibilities for foodbased therapies. Eur J Pharmacol 817: 86-95. https://doi.org/10.1016/j.ejphar.2017.05.042
|
| [67] |
Kim S, Kwon SH, Kam TI, et al. (2019) Transneuronal propagation ff pathologic α-synuclein from the gut to the brain models Parkinson's disease. Neuron 103: 627-641. https://doi.org/10.1016/j.neuron.2019.05.035
|
| [68] |
Sampson TR, Debelius JW, Thron T, et al. (2016) Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson's disease. Cell 167: 1469-1480. https://doi.org/10.1016/j.cell.2016.11.018
|
| [69] |
Sun MF, Shen YQ (2018) Dysbiosis of gut microbiota and microbial metabolites in Parkinson's disease. Ageing Res Rev 45: 53-61. https://doi.org/10.1016/j.arr.2018.04.004
|
| [70] |
Marizzoni M, Provasi S, Cattaneo A, et al. (2017) Microbiota and neurodegenerative diseases. Curr Opin Neurol 30: 630-638. https://doi.org/10.1097/WCO.0000000000000496
|
| [71] |
Perez-Pardo P, Kliest T, Dodiya HB, et al. (2017) The gut-brain axis in Parkinson's disease: Possibilities for foodbased therapies. Eur J Pharmacol 817: 86-95. https://doi.org/10.1016/j.ejphar.2017.05.042
|
| [72] |
van den Hoogen WJ, Laman JD, 't Hart BA (2017) Modulation of multiple sclerosis and its animal model experimental autoimmune encephalomyelitis by food and gut microbiota. Front Immunol 8: 1081. https://doi.org/10.3389/fimmu.2017.01081
|
| [73] |
Mowry EM, Glenn JD (2018) The dynamics of the gut microbiome in multiple sclerosis in relation to disease. Neurol Clin 36: 185-196. https://doi.org/10.1016/j.ncl.2017.08.008
|
| [74] | Chu F, Shi M, Lang Y, et al. (2018) Gut microbiota in multiple sclerosis and experimental autoimmune encephalomyelitis: current applications and future perspectives. Mediators Inflamm 2018: 8168717. https://doi.org/10.1155/2018/8168717 |
| [75] |
Jiang HY, Zhang X, Yu ZH, et al. (2018) Altered gut microbiota profile in patients with generalized anxiety disorder. J Psychiatr Res 104: 130-136. https://doi.org/10.1016/j.jpsychires.2018.07.007
|
| [76] |
Rogers GB, Keating DJ, Young RL, et al. (2016) From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol Psychiatr 21: 738-748. https://doi.org/10.1038/mp.2016.50
|
| [77] |
Tognini P (2017) Gut microbiota: A potential regulator of neurodevelopment. Front Cell Neurosci 11: 25. https://doi.org/10.3389/fncel.2017.00025
|
| [78] |
Valles-Colomer M, Falony G, Darzi Y, et al. (2019) The neuroactive potential of the human gut microbiota in quality of life and depression. Nat Microbiol 4: 623-632. https://doi.org/10.1038/s41564-018-0337-x
|
| [79] |
Dash S, Syed YA, Khan MR (2022) Understanding the Role of the Gut Microbiome in Brain Development and Its Association With Neurodevelopmental Psychiatric Disorders. Front Cell Dev Biol 10: 880544. https://doi.org/10.3389/fcell.2022.880544
|
| [80] |
Frémont M, Coomans D, Massart S, et al. (2013) High-throughput 16S rRNA Gene Sequencing Reveals Alterations of Intestinal Microbiota in Myalgic Encephalomyelitis/chronic Fatigue Syndrome Patients. Anaerobe 22. https://doi.org/10.1016/j.anaerobe.2013.06.002
|
| [81] |
Skonieczna-Żydecka K, Łoniewski I, Maciejewska D, et al. (2017) Mikrobiota jelitowa i składniki pokarmowe jako determinanty funkcji układu nerwowego. Część I. Mikrobiota przewodu pokarmowego. Aktual Neurol 17: 181-188. https://doi.org/10.15557/AN.2017.0020
|
| [82] |
Strati F, Cavalieri D, Albanese D, et al. (2017) New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 5: 24. https://doi.org/10.1186/s40168-017-0242-1
|
| [83] |
Hashimoto K (2023) Emerging role of the host microbiome in neuropsychiatric disorders: overview and future directions. Mol Psychiatr 28: 3625-3637. https://doi.org/10.1038/s41380-023-02287-6
|
| [84] |
Hou K, Wu ZX, Chen XY, et al. (2022) Microbiota in health and disease. Signal Transduct Target Ther 7: 135. https://doi.org/10.1038/s41392-022-00974-4
|
| [85] |
Aggarwal N, Kitano S, Puah GRY, et al. (2023) Microbiome and human health: current understanding, engineering, and enabling technologies. Chem Rev 123: 31-72. https://doi.org/10.1021/acs.chemrev.2c00431
|
| [86] |
Martínez M, Postolache TT, García-Bueno B, et al. (2022) The role of the oral microbiota related to periodontal diseases in anxiety, mood and trauma- and stress-related disorders. Front Psych 12: 814177. https://doi.org/10.3389/fpsyt.2021.814177
|
| [87] |
Kisely S, Sawyer E, Siskind D, et al. (2016) The oral health of people with anxiety and depressive disorders-a systematic review and meta-analysis. J Affect Disord 200: 119-32. https://doi.org/10.1016/j.jad.2016.04.040
|
| [88] |
Coelho JMF, Miranda SS, da Cruz SS, et al. (2020) Common mental disorder is associated with periodontitis. J Periodontal Res 55: 221-8. https://doi.org/10.1111/jre.12705
|
| [89] |
Dominy SS, Lynch C, Ermini F, et al. (2019) Porphyromonas gingivalis in Alzheimer's disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci Adv 5: eaau3333. https://doi.org/10.1126/sciadv.aau3333
|
| [90] |
Pisani F, Pisani V, Arcangeli F, et al. (2023) Treponema denticola Has the Potential to Cause Neurodegeneration in the Midbrain via the Periodontal Route of Infection-Narrative Review. Int J Environ Res Public Health 20: 6049. https://doi.org/10.3390/ijerph20116049
|
| [91] |
Emery DC, Shoemark DK, Batstone TE, et al. (2017) 16S rRNA next generation sequencing analysis shows bacteria in Alzheimer's post-mortem brain. Front Aging Neurosci 9: 195. https://doi.org/10.3389/fnagi.2017.00195
|
| [92] |
Siddiqui H, Rribe ERK, Singhrao SK, et al. (2019) High throughput sequencing detects gingivitis and periodontal oral bacteria in Alzheimer's disease autopsy brains. Neuro Res 1: 3. https://doi.org/10.35702/nrj.10003
|
| [93] |
Weber C, Dilthey A, Finzer P (2023) The role of microbiome–host interactions in the development of Alzheimer's disease. Front Cell Infect Microbiol 13: 1151021. https://doi.org/10.3389/fcimb.2023.1151021
|
| [94] |
Emery DC, Davies M, Cerajewska TL, et al. (2022) High resolution 16S rRNA gene Next Generation Sequencing study of brain areas associated with Alzheimer's and Parkinson's disease. Front Aging Neurosci 14: 1026260. https://doi.org/10.3389/fnagi.2022.1026260
|
| [95] |
Berthouzoz E, Lazarevic V, Zekeridou A, et al. (2023) Oral and intestinal dysbiosis in Parkinson's disease. Rev Neurol (Paris) 179: 937-946. https://doi.org/10.1016/j.neurol.2022.12.010
|
| [96] |
Jo S, Kang W, Hwang YS, et al. (2022) Oral and gut dysbiosis leads to functional alterations in Parkinson's disease. NPJ Parkinsons Dis 8: 87. https://doi.org/10.1038/s41531-022-00351-6
|
| [97] |
Li Z, Lu G, Luo E, et al. (2022) Oral, nasal, and gut microbiota in Parkinson's disease. Neuroscience 480: 65-78. https://doi.org/10.1016/j.neuroscience.2021.10.011
|
| [98] |
Goswami A, Wendt FR, Pathak GA, et al. (2021) Role of microbes in the pathogenesis of neuropsychiatric disorders. Front Neuroendocrinol 21: 100917. https://doi.org/10.1016/j.yfrne.2021.100917
|
| [99] |
Ferraretto A, Donetti E, García-Mena J, et al. (2023) Editorial: The gut-skin-brain axis in human health and disease. Front Nutr 10: 1155614. https://doi.org/10.3389/fnut.2023.1155614
|
| [100] |
Hermes BM, Rademacher F, Chung C, et al. (2022) Skin microbiota analysis in patients with anorexia nervosa and healthy-weight controls reveals microbial indicators of healthy weight and associations with the antimicrobial peptide psoriasin. Sci Rep 12: 15515. https://doi.org/10.1038/s41598-022-19676-6
|
| [101] |
Arikan M, Yildiz Z, Kahraman Demir T, et al. (2022) Axillary microbiota is associated with cognitive impairment in Parkinson's disease patients. Microbiol Spectr 10: e0235821. https://doi.org/10.1128/spectrum.02358-21
|
| [102] |
Armstrong AW, Read C (2020) Pathophysiology, clinical presentation, and treatment of psoriasis: a review. JAMA 323: 1945-60. https://doi.org/10.1001/jama.2020.4006
|
| [103] |
Griffiths CEM, Armstrong AW, Gudjonsson JE, et al. (2021) Psoriasis. Lancet 397: 1301-15. https://doi.org/10.1016/S0140-6736(20)32549-6
|
| [104] |
Hadian Y, Fregoso D, Nguyen C, et al. (2020) Microbiome-skin-brain axis: A novel paradigm for cutaneous wounds. Wound Repair Regen 28: 282-92. https://doi.org/10.1111/wrr.12800
|
| [105] |
Woo YR, Han YJ, Kim HS, et al. (2020) Updates on the risk of neuropsychiatric and gastrointestinal comorbidities in rosacea and its possible relationship with the gut-brain-skin axis. Int J Mol Sci 21: 8427. https://doi.org/10.3390/ijms21228427
|
| [106] |
Chen G, Chen ZM, Fan XY, et al. (2021) Gut-brain-skin axis in psoriasis: a review. Dermatol Ther (Heidelb) 11: 25-38. https://doi.org/10.1007/s13555-020-00466-9
|
| [107] |
Wang X, Li Y, Wu L, et al. (2021) Dysregulation of the gut-brain-skin axis and key overlapping inflammatory and immune mechanisms of psoriasis and depression. Biomed Pharmacother 137: 111065. https://doi.org/10.1016/j.biopha.2020.111065
|
| [108] |
Shinno-Hashimoto H, Hashimoto Y, Wei Y, et al. (2021) Abnormal composition of microbiota in the gut and skin of imiquimod-treated mice. Sci Rep 11: 112665. https://doi.org/10.1038/s41598-021-90480-4
|
| [109] |
Kumpitsch C, Koskinen K, Schöpf V, et al. (2019) The microbiome of the upper respiratory tract in health and disease. BMC Biol 17: 87. https://doi.org/10.1186/s12915-019-0703-z
|
| [110] |
Tai J, Han MS, Kwak J, et al. (2021) Association between microbiota and nasal mucosal diseases in terms of immunity. Int J Mol Sci 22: 4744. https://doi.org/10.3390/ijms22094744
|
| [111] |
Thangaleela S, Sivamaruthi BS, Kesika P, et al. (2022) Nasal microbiota, olfactory health, neurological disorders and aging—a review. Microorganisms 10: 1405. https://doi.org/10.3390/microorganisms10071405
|
| [112] |
Xie J, Tian S, Liu J, et al. (2022) Dual role of the nasal microbiota in neurological diseases-An unignorable risk factor or a potential therapy carrier. Pharmacol Res 179: 106189. https://doi.org/10.1016/j.phrs.2022.106189
|
| [113] |
Sarkar A, Lehto SM, Harty S, et al. (2016) Psychobiotics and the manipulation of bacteria-gut-brain signals. Trends Neurosci 39: 763-781. https://doi.org/10.1016/j.tins.2016.09.002
|
| [114] |
Snigdha S, Ha K, Tsai P, et al. (2022) Probiotics: Potential novel therapeutics for microbiota-gut-brain axis dysfunction across gender and lifespan. Pharmacol Ther 231: 107978. https://doi.org/10.1016/j.pharmthera.2021.107978
|
| [115] |
Chung Y, Jin H, Cui Y (2014) Fermented milk of Lactobacillus helveticus IDCC3801 improves cognitive functioning during cognitive fatigue tests in healthy older adults. J Funct Foods 10: 465-474. https://doi.org/10.1016/j.jff.2014.07.007
|
| [116] | Allen AP, Hutch W, Borre YE, et al. (2016) Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Translat. Psychiatry 6: 939-939. https://doi.org/10.1038/tp.2016.191 |
| [117] |
Moloney GM, Long-Smith CM, Murphy A (2020) Improvements in sleep indices during exam stress due to consumption of a Bifidobacterium longum. Brain Behav Immun Health 10: 100174. https://doi.org/10.1016/j.bbih.2020.100174
|
| [118] |
Zhao M, Ren Z, Zhao A, et al. (2024) Gut bacteria-driven homovanillic acid alleviates depression by modulating synaptic integrity. Cell Metab 36: 1000-1012.e6. https://doi.org/10.1016/j.cmet.2024.03.010
|
| [119] |
Chong HX, Yusoff NAA, Hor YY, et al. (2019) Lactobacillus plantarum DR7 alleviates stress and anxiety in adults: a randomised, double-blind, placebo-cont controlled study. Benef Microbes 10: 355-373. https://doi.org/10.3920/BM2018.0135
|
| [120] |
Kelly JR, Allen AP, Temko A, et al. (2017) Lost in translation? The potential psychobiotic Lactobacillus rhamnosus (JB-1) fails to modulate stress or cognitive performance in healthy male subjects. Brain Behav Immun 61: 50-59. https://doi.org/10.1016/j.bbi.2016.11.018
|
| [121] |
Kochalska K, Oakden W, Słowik T, et al. (2020) Dietary supplementation with Lactobacillus rhamnosus JB-1 restores brain neurochemical balance and mitigates the progression of mood disorder in a rat model of chronic unpredictable mild stress. Nutr Res 82: 44-57. https://doi.org/10.1016/j.nutres.2020.06.019
|
| [122] |
Jarosz ŁS, Socała K, Michalak K, et al. (2024) The effect of psychoactive bacteria, Bifidobacterium longum Rosell®-175 and Lactobacillus rhamnosus JB-1, on brain proteome profiles in mice. Psychopharmacology (Berl) 241: 925-945. https://doi.org/10.1007/s00213-023-06519-z
|
| [123] |
Yunes RA, Poluektova EU, Vasileva EV, et al. (2020) A Multi-strain Potential Probiotic Formulation of GABA-Producing Lactobacillus plantarum 90sk and Bifidobacterium adolescentis 150 with Antidepressant Effects. Probiotics Antimicrob. Proteins 12: 973-979. https://doi.org/10.1007/s12602-019-09601-1
|
| [124] |
De Oliveira FL, Salgaço MK, de Oliveira MT, et al. (2023) Exploring the Potential of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 as Promising Psychobiotics Using SHIME. Nutrients 15: 1521. https://doi.org/10.3390/nu15061521
|
| [125] |
Asaoka D, Xiao J, Takeda T, et al. (2022) Effect of Probiotic Bifidobacterium breve in Improving Cognitive Function and Preventing Brain Atrophy in Older Patients with Suspected Mild Cognitive Impairment: Results of a 24-Week Randomized, Double-Blind, Placebo-Controlled Trial. J Alzheimers Dis 88: 75-95. https://doi.org/10.3233/JAD-220148
|
Figures(1) / Tables(1)

mgr Natalia Białoń, dr hab. n. o zdr. Dariusz Górka, mgr Mikołaj Górka. The brain-gut axis: communication mechanisms and the role of the microbiome as a neuroprotective factor in the development of neurodegenerative diseases: A literature overview[J]. AIMS Neuroscience, 2024, 11(3): 289-311. doi: 10.3934/Neuroscience.2024019







 DownLoad:
DownLoad: