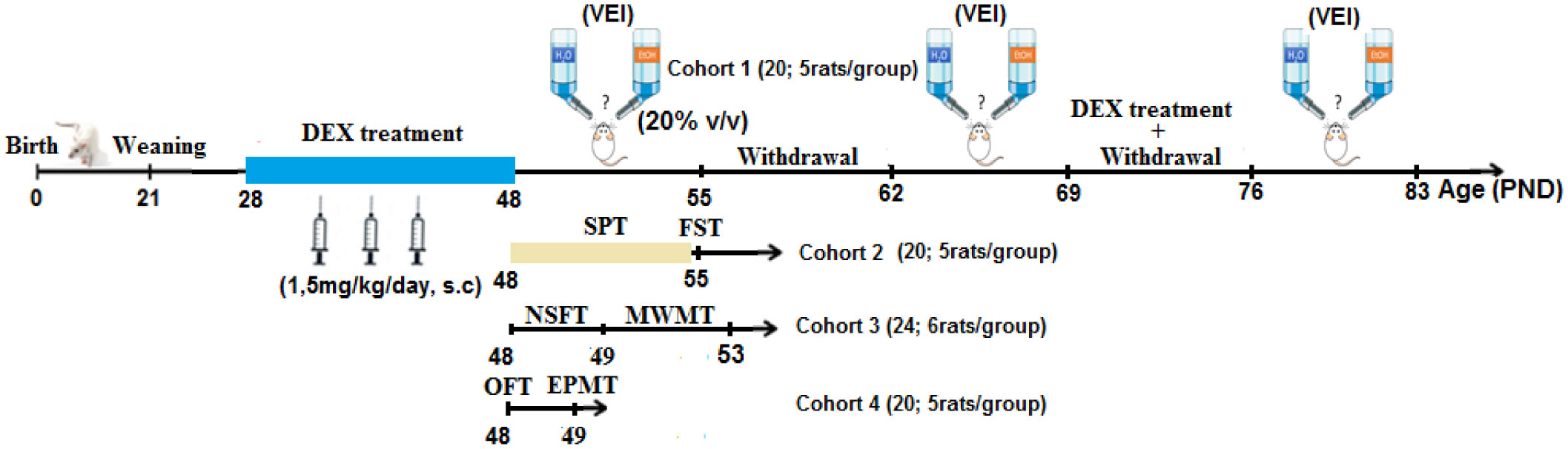
The stress response is attached to psychosomatic and psychiatric disorders. Therefore, it is important to comprehend the underlying mechanisms influencing this relationship. Moreover, men and women respond differently to stress–both psychologically and biologically. These differences should be studied to have an enhanced understanding of the gender difference. However, researches shedding light on sex dimorphism implication have historically been insufficient. Based on observations that advocate the inclusion of sex as a biological variable in stress response, the present study was designed to explore sex differences in (i) depressive-like, (ii) anxiety-like behaviors, (iii) cognitive-like performances, and (iv) voluntary ethanol intake (VEI) in Wistar rat submitted to dexamethasone (DEX)-stress simulation. Rats were administered daily with DEX (1.5 mg/kg, s.c., 21 days) or vehicle. Behavior, cognitive, and VEI states of rats were evaluated in the following paradigms: forced swimming test (FST); saccharin preference test (SPT); open field test (OFT); elevated plus-maze test (EPMT); novelty suppressed feeding test (NSFT); spatial learning and memory in Morris water maze test (MWMT); VEI in two-bottle choice paradigm. DEX-treated rats showed a set of depression-like behaviors: increased time of immobility; reduced preference for saccharin consumption; increased anxiety-like behavior; cognitive impairments; and enhanced VEI. Sexual dimorphism was recorded in this study. Females were more impaired in FST, SPT, EPMT, NSFT, and VEI. Results demonstrate that DEX-treatment induced a behavioral alterations related to anxiety-depressive-like state with learning and memory impairments; confirm the facilitatory role of glucocorticoids on VEI and reveal sexual dimorphism in stress response.
Citation: Laaziz Abderrahim, El Mostafi Hicham, Elhessni Aboubaker, Azeroil Fatima, Touil Tarik, Boumlah Soufiane, Mesfioui Abdelhalim. Sex differences in behavioral, cognitive and voluntary ethanol-intake effects in Dexamethasone-induced depression-like state in Wistar rat[J]. AIMS Neuroscience, 2022, 9(2): 228-249. doi: 10.3934/Neuroscience.2022012
The stress response is attached to psychosomatic and psychiatric disorders. Therefore, it is important to comprehend the underlying mechanisms influencing this relationship. Moreover, men and women respond differently to stress–both psychologically and biologically. These differences should be studied to have an enhanced understanding of the gender difference. However, researches shedding light on sex dimorphism implication have historically been insufficient. Based on observations that advocate the inclusion of sex as a biological variable in stress response, the present study was designed to explore sex differences in (i) depressive-like, (ii) anxiety-like behaviors, (iii) cognitive-like performances, and (iv) voluntary ethanol intake (VEI) in Wistar rat submitted to dexamethasone (DEX)-stress simulation. Rats were administered daily with DEX (1.5 mg/kg, s.c., 21 days) or vehicle. Behavior, cognitive, and VEI states of rats were evaluated in the following paradigms: forced swimming test (FST); saccharin preference test (SPT); open field test (OFT); elevated plus-maze test (EPMT); novelty suppressed feeding test (NSFT); spatial learning and memory in Morris water maze test (MWMT); VEI in two-bottle choice paradigm. DEX-treated rats showed a set of depression-like behaviors: increased time of immobility; reduced preference for saccharin consumption; increased anxiety-like behavior; cognitive impairments; and enhanced VEI. Sexual dimorphism was recorded in this study. Females were more impaired in FST, SPT, EPMT, NSFT, and VEI. Results demonstrate that DEX-treatment induced a behavioral alterations related to anxiety-depressive-like state with learning and memory impairments; confirm the facilitatory role of glucocorticoids on VEI and reveal sexual dimorphism in stress response.

| [1] | Arango-Lievano M, Kaplitt MG (2015) entre la dépression et l'addiction. méd/sci 31: 546-50. https://doi.org/10.1051/medsci/20153105017 |
| [2] |
Herman JP, Cullinan WE (1997) Neurocircuitry of stress: central control of the hypothalamo–pituitary–adrenocortical axis. Trends neurosci 20: 78-84. https://doi.org/10.1016/S0166-2236(96)10069-2 
|
| [3] |
Tsigos C, Chrousos GP (2002) Hypothalamic–pituitary–adrenal axis, neuroendocrine factors and stress. J Psychosom Res 53: 865-871. https://doi.org/10.1016/S0022-3999(02)00429-4
|
| [4] |
McEwen BS (2000) Effects of adverse experiences for brain structure and function. Biol Psychiatry 48: 721-731. https://doi.org/10.1016/S0006-3223(00)00964-1
|
| [5] |
David DJ, Samuels BA, Rainer Q, et al. (2009) Neurogenesis-dependent and-independent effects of fluoxetine in an animal model of anxiety/depression. Neuron 62: 479-493. https://doi.org/10.1016/j.neuron.2009.04.017
|
| [6] |
Sapolsky RM, Krey LC, McEwen BS (1984) Glucocorticoid-sensitive hippocampal neurons are involved in terminating the adrenocortical stress response. Proc Natl Acad Sci 81: 6174-6177. https://doi.org/10.1073/pnas.81.19.6174
|
| [7] |
Sapolsky RM, Krey LC, McEWEN BS (1985) Prolonged glucocorticoid exposure reduces hippocampal neuron number: implications for aging. J Neurosci 5: 1222-1227. https://doi.org/10.1523/JNEUROSCI.05-05-01222.1985
|
| [8] |
Vyas A, Bernal S, Chattarji S (2003) Effects of chronic stress on dendritic arborization in the central and extended amygdala. Brain Res 965: 290-294. https://doi.org/10.1016/S0006-8993(02)04162-8
|
| [9] |
Pham K, Nacher J, Hof PR, et al. (2003) Repeated restraint stress suppresses neurogenesis and induces biphasic PSA-NCAM expression in the adult rat dentate gyrus. Eur J Neurosci 17: 879-886. https://doi.org/10.1046/j.1460-9568.2003.02513.x
|
| [10] |
Sterner EY, Kalynchuk LE (2010) Behavioral and neurobiological consequences of prolonged glucocorticoid exposure in rats: relevance to depression. Prog Neuro-Psychopharmacology Biol Psychiatry 34: 777-790. https://doi.org/10.1016/j.pnpbp.2010.03.005
|
| [11] |
Parker KJ, Schatzberg AF, Lyons DM (2003) Neuroendocrine aspects of hypercortisolism in major depression. Horm Behav 43: 60-66. https://doi.org/10.1016/S0018-506X(02)00016-8
|
| [12] | MacQueen GM, Campbell S, McEwen BS, et al. (2005) Course of illness, hippocampal function, and hippocampal volume in major depression. Focus 100: 1387-155. https://doi.org/10.1176/foc.3.1.146 |
| [13] |
Alcalar N, Ozkan S, Kadioglu P, et al. (2013) Evaluation of depression, quality of life and body image in patients with Cushing's disease. Pituitary 16: 333-340. https://doi.org/10.1007/s11102-012-0425-5
|
| [14] |
Lamers F, van Oppen P., Comijs HC, et al. (2011) Comorbidity patterns of anxiety and depressive disorders in a large cohort study: the Netherlands Study of Depression and Anxiety (NESDA). J Clin Psychiatry 72: 3397. https://doi.org/10.4088/JCP.10m06176blu
|
| [15] |
Blaine SK., Sinha R (2017) Alcohol, stress, and glucocorticoids: From risk to dependence and relapse in alcohol use disorders. Neuropharmacology 122: 136-147. https://doi.org/10.1016/j.neuropharm.2017.01.037
|
| [16] |
Fahlke C, Eriksson CP (2000) Effect of adrenalectomy and exposure to corticosterone on alcohol intake in alcohol-preferring and alcohol-avoiding rat lines. Alcohol Alcohol 35: 139-144. https://doi.org/10.1093/alcalc/35.2.139
|
| [17] |
Fahlke C, Hansen S (1996) Facilitation of ethanol consumption by intracerebroventricular infusions of corticosterone. Psychopharmacol 127: 133-139. https://doi.org/10.1007/BF02805986
|
| [18] |
Piazza PV, Deroche V, Deminiere JM, et al. (1993) Corticosterone in the range of stress-induced levels possesses reinforcing properties: implications for sensation-seeking behaviors. Proc Natl Acad Sci 90: 11738-11742. https://doi.org/10.1073/pnas.90.24.11738
|
| [19] |
Brand I, Fliegel S, Spanagel R, et al. (2013) Global ethanol-induced enhancements of monoaminergic neurotransmission: a meta-analysis study. Alcohol: Clin Exp Res 37: 2048-2057. https://doi.org/10.1111/acer.12207
|
| [20] |
Spanagel R, Noori HR, Heilig M (2014) Stress and alcohol interactions: animal studies and clinical significance. Trends Neurosci 37: 219-227. https://doi.org/10.1016/j.tins.2014.02.006
|
| [21] |
Beery AK, Zucker I (2011) Sex bias in neuroscience and biomedical research. Neurosci Biobeha Rev 35: 565-572. https://doi.org/10.1016/j.neubiorev.2010.07.002
|
| [22] |
Shors TJ, Chua C, Falduto J (2001) Sex differences and opposite effects of stress on dendritic spine density in the male versus female hippocampus. J Neurosci 21: 6292-6297. https://doi.org/10.1523/JNEUROSCI.21-16-06292.2001
|
| [23] |
Cosgrove KP, Mazure CM, Staley JK (2007) Evolving knowledge of sex differences in brain structure, function, and chemistry. Biol Psychiatry 62: 847-855. https://doi.org/10.1016/j.biopsych.2007.03.001
|
| [24] |
Lind KE., Gutierrez EJ, Yamamoto DJ, et al. (2017) Sex disparities in substance abuse research: Evaluating 23 years of structural neuroimaging studies. Drug Alcohol Depend 173: 92-98. https://doi.org/10.1016/j.drugalcdep.2016.12.019
|
| [25] |
Miller LR, Marks C, Becker JB, et al. (2017) Considering sex as a biological variable in preclinical research. FASEB J 31: 29-34. https://doi.org/10.1096/fj.201600781r
|
| [26] |
Choleris E, Galea LA, Sohrabji F, et al. (2018) Sex differences in the brain: Implications for behavioral and biomedical research. Neurosci Biobeha Rev 85: 126-145. https://doi.org/10.1016/j.neubiorev.2017.07.005
|
| [27] |
Clayton JA, Collins FS (2014) Policy: NIH to balance sex in cell and animal studies. Nat News 509: 282. https://doi.org/10.1038/509282a
|
| [28] |
Guizzetti M, Davies DL, Egli M, et al. (2016) Sex and the lab: an alcohol-focused commentary on the NIH initiative to balance sex in cell and animal studies. Alcohol: Clin Exp Res 40: 1182-1191. https://doi.org/10.1111/acer.13072
|
| [29] |
Sigwalt AR, Budde H, Helmich I, et al. (2011) Molecular aspects involved in swimming exercise training reducing anhedonia in a rat model of depression. Neurosci 192: 661-674. https://doi.org/10.1016/j.neuroscience.2011.05.075
|
| [30] |
Kenna HA, Poon AW, de los Angeles CP, et al. (2011) Psychiatric complications of treatment with corticosteroids: review with case report. Psychiatry clin Neurosci 65: 549-560. https://doi.org/10.1111/j.1440-1819.2011.02260.x
|
| [31] |
Gaspar R, Soares-Cunha C, Domingues AV, et al. (2021) Resilience to stress and sex-specific remodeling of microglia and neuronal morphology in a rat model of anxiety and anhedonia. Neurobiol Stress 14: 100302. https://doi.org/10.1016/j.ynstr.2021.100302
|
| [32] |
Skupio U, Tertil M, Sikora M, et al. (2015) Behavioral and molecular alterations in mice resulting from chronic treatment with dexamethasone: relevance to depression. Neurosci 286: 141-150. https://doi.org/10.1016/j.neuroscience.2014.11.035
|
| [33] |
Ferreira AS, Galvão S, Gaspar R, Rodrigues-Neves AC, et al. (2021) Sex-specific changes in peripheral metabolism in a model of chronic anxiety induced by prenatal stress. Eur J Clin Invest 51: e13639. https://doi.org/10.1111/eci.13639
|
| [34] |
Mesripour A, Alhimma F, Hajhashemi V (2019) The effect of vitamin B6 on dexamethasone-induced depression in mice model of despair. Nutr Neurosci 22: 744-749. https://doi.org/10.1080/1028415X.2018.1442184
|
| [35] |
Iacono LL, Carola V (2018) The impact of adolescent stress experiences on neurobiological development. Semi cell deve biol 77: 93-103. https://doi.org/10.1016/j.semcdb.2017.09.040
|
| [36] |
Yohn NL, Blendy JA (2017) Adolescent chronic unpredictable stress exposure is a sensitive window for long-term changes in adult behavior in mice. Neuropsychopharmacology 42: 1670-1678. https://doi.org/10.1038/npp.2017.11
|
| [37] |
Porsolt RD, Le Pichon M, Jalfre ML (1977) Depression: a new animal model sensitive to antidepressant treatments. Nat 266: 730-732. https://doi.org/10.1038/266730a0
|
| [38] |
Abelaira HM, Réus GZ, Quevedo J (2013) Animal models as tools to study the pathophysiology of depression. Braz J Psychiatry 35: S112-S120. https://doi.org/10.1590/1516-4446-2013-1098
|
| [39] |
Hazra S, Kumar S, Saha GK, et al. (2015) Chronic Administration of Bacopa monniera Alleviates Depressive Like Behavior and Increases the Expression of ERK1/2 in Hippocampus and Pre-Frontal Cortex of Chronic Unpredictable Stress Induced Rats. Int Neuropsychiatr Dis J 3: 47-58. 10.9734/INDJ/2015/14414
|
| [40] |
Clénet F, Bouyon E, Hascoët M, et al. (2006) Light/dark cycle manipulation influences mice behaviour in the elevated plus maze. Behav Brain Res 166: 140-149. https://doi.org/10.1016/j.bbr.2005.07.018
|
| [41] |
Francis-Oliveira J, Ponte B, Barbosa APM, et al. (2013) Fluoxetine exposure during pregnancy and lactation: Effects on acute stress response and behavior in the novelty-suppressed feeding are age and gender-dependent in rats. Behav Brain Res 252: 195-203. https://doi.org/10.1016/j.bbr.2013.05.064
|
| [42] | Wenk GL (2004) Assessment of spatial memory using the radial arm maze and Morris water maze. Curr Prot Neurosci 26: 8.5 A. 1-8.5 A. 12. https://doi.org/10.1002/0471142301.ns0805as26 |
| [43] |
Monfort P, Erceg S, Piedrafita B, et al. (2007) Chronic liver failure in rats impairs glutamatergic synaptic transmission and long-term potentiation in hippocampus and learning ability. Eur J Neurosci 25: 2103-2111. https://doi.org/10.1111/j.1460-9568.2007.05444.x
|
| [44] |
El Mostafi H, Elhessni A, Touil T, et al. (2020) Argan oil supplementation attenuates voluntary ethanol consumption and withdrawal syndrome promoted by adolescent intermittent ethanol in rat. Alcohol 87: 39-50. https://doi.org/10.1016/j.alcohol.2020.04.007
|
| [45] | Peñasco S, Mela V, López-Moreno JA, et al. (2015) Early maternal deprivation enhances voluntary alcohol intake induced by exposure to stressful events later in life. Neural Plast . https://doi.org/10.1155/2015/342761 |
| [46] |
Ruksee N, Tongjaroenbuangam W, Mahanam T, et al. (2014) Melatonin pretreatment prevented the effect of dexamethasone negative alterations on behavior and hippocampal neurogenesis in the mouse brain. J Steroid Bbiochem Mol Biol 143: 72-80. https://doi.org/10.1016/j.jsbmb.2014.02.011
|
| [47] |
Lebedeva KA, Caruncho HJ, Kalynchuk LE (2017) Cyclical corticosterone administration sensitizes depression-like behavior in rats. Neurosci Lett 650: 45-51. https://doi.org/10.1016/j.neulet.2017.04.023
|
| [48] |
Ali SH, Madhana RM, Athira KV, et al. (2015) Resveratrol ameliorates depressive-like behavior in repeated corticosterone-induced depression in mice. Steroids 101: 37-42. https://doi.org/10.1016/j.steroids.2015.05.010
|
| [49] |
Luo GQ, Liu L, Gao QW, et al. (2017) Mangiferin prevents corticosterone-induced behavioural deficits via alleviation of oxido-nitrosative stress and down-regulation of indoleamine 2, 3-dioxygenase (IDO) activity. Neurol Res 39: 709-718. https://doi.org/10.1080/01616412.2017.1310705
|
| [50] |
Bai Y, Song L, Dai G, et al. (2018) Antidepressant effects of magnolol in a mouse model of depression induced by chronic corticosterone injection. Steroids 135: 73-78. https://doi.org/10.1016/j.steroids.2018.03.005
|
| [51] |
Xu J, Wang R, Liu Y, et al. (2019) Short-and long-term alterations of FKBP5-GR and specific microRNAs in the prefrontal cortex and hippocampus of male rats induced by adolescent stress contribute to depression susceptibility. Psychoneuroendocrinology 101: 204-215. https://doi.org/10.1016/j.psyneuen.2018.11.008
|
| [52] |
Hill MN, Brotto LA, Lee TTY, et al. (2003) Corticosterone attenuates the antidepressant-like effects elicited by melatonin in the forced swim test in both male and female rats. Prog Neuro-Psychopharmacol Biol Psychiatry 27: 905-911. https://doi.org/10.1016/S0278-5846(03)00149-0
|
| [53] |
Brotto LA, Barr AM, Gorzalka BB (2000) Sex differences in forced-swim and open-field test behaviours after chronic administration of melatonin. Eur J Pharmacol 402: 87-93. https://doi.org/10.1016/S0014-2999(00)00491-X
|
| [54] |
Wintink AJ, Young NA, Davis AC, et al. (2003) Kindling-induced emotional behavior in male and female rats. Behav Neurosci 117: 632. https://doi.org/10.1037/0735-7044.117.3.632
|
| [55] |
Alonso SJ, Castellano MA, Afonso D, et al. (1991) Sex differences in behavioral despair: relationships between behavioral despair and open field activity. Physiol behavi 49: 69-72. https://doi.org/10.1016/0031-9384(91)90232-D
|
| [56] |
Johnson SA, Fournier NM, Kalynchuk LE (2006) Effect of different doses of corticosterone on depression-like behavior and HPA axis responses to a novel stressor. Behav Brain Res 168: 280-288. https://doi.org/10.1016/j.bbr.2005.11.019
|
| [57] |
Feng Y, Rhodes PG, Liu H, et al. (2009) Dexamethasone induces neurodegeneration but also up-regulates vascular endothelial growth factor A in neonatal rat brains. Neurosci 158: 823-832. https://doi.org/10.1016/j.neuroscience.2008.10.024
|
| [58] |
Brummelte S, Pawluski JL, Galea LA (2006) High post-partum levels of corticosterone given to dams influence postnatal hippocampal cell proliferation and behavior of offspring: a model of post-partum stress and possible depression. Horm Behave 50: 370-382. https://doi.org/10.1016/j.yhbeh.2006.04.008
|
| [59] |
Kalynchuk LE, Gregus A, Boudreau D, et al. (2004) Corticosterone increases depression-like behavior, with some effects on predator odor-induced defensive behavior, in male and female rats. Behav Neurosci 118: 1365. https://doi.org/10.1037/0735-7044.118.6.1365
|
| [60] |
Casarotto PC, Andreatini R (2007) Repeated paroxetine treatment reverses anhedonia induced in rats by chronic mild stress or dexamethasone. Eur Neuropsychopharmacology 17: 735-742. https://doi.org/10.1016/j.euroneuro.2007.03.001
|
| [61] |
Page GG, Opp MR, Kozachik SL (2016) Sex differences in sleep, anhedonia, and HPA axis activity in a rat model of chronic social defeat. Neurobiol Stress 3: 105-113. https://doi.org/10.1016/j.ynstr.2016.03.002
|
| [62] |
Kenna HA, Poon AW, de los Angeles CP, et al. (2011) Psychiatric complications of treatment with corticosteroids: review with case report. Psychiatry Clin Neurosci 65: 549-560. https://doi.org/10.1111/j.1440-1819.2011.02260.x
|
| [63] |
Skórzewska A, Bidziński A, Lehner M, et al. (2006) The effects of acute and chronic administration of corticosterone on rat behavior in two models of fear responses, plasma corticosterone concentration, and c-Fos expression in the brain structures. Pharmacol Biochem Behav 85: 522-534. https://doi.org/10.1016/j.pbb.2006.10.001
|
| [64] |
Lee B, Shim I, Lee HJ, Yang Y, et al. (2009) Effects of acupuncture on chronic corticosterone-induced depression-like behavior and expression of neuropeptide Y in the rats. Neurosci lett 453: 151-156. https://doi.org/10.1016/j.neulet.2009.01.076
|
| [65] |
Johnston AL, File SE (1991) Sex differences in animal tests of anxiety. Physiol behave 49: 245-250. https://doi.org/10.1016/0031-9384(91)90039-Q
|
| [66] | Sandi C, Pinelo-Nava MT (2007) Stress and memory: behavioral effects and neurobiological mechanisms. Neural Plast . https://doi.org/10.1155/2007/78970 |
| [67] |
Lupien SJ, McEwen BS (1997) The acute effects of corticosteroids on cognition: integration of animal and human model studies. Brain Res Rev 24: 1-27. https://doi.org/10.1016/S0165-0173(97)00004-0
|
| [68] |
Newcomer JW, Selke G, Melson AK, et al. (1999) Decreased memory performance in healthy humans induced by stress-level cortisol treatment. Arch Gen Psychiatry 56: 527-533. 10-1001/pubs.Arch Gen Psychiatry-ISSN-0003-990x-56-6-yoa8291
|
| [69] |
Sapolsky RM (2000) The possibility of neurotoxicity in the hippocampus in major depression: a primer on neuron death. Biol Psychiatry 48: 755-765. https://doi.org/10.1016/S0006-3223(00)00971-9
|
| [70] |
Patki G, Solanki N, Atrooz F, et al. (2013) Depression, anxiety-like behavior and memory impairment are associated with increased oxidative stress and inflammation in a rat model of social stress. Brain Res 1539: 73-86. https://doi.org/10.1016/j.brainres.2013.09.033
|
| [71] |
Richter SH, Zeuch B, Lankisch K, et al. (2013) Where have I been? Where should I go? Spatial working memory on a radial arm maze in a rat model of depression. PloS One 8: e62458. https://doi.org/10.1371/journal.pone.0062458
|
| [72] |
Song L, Che W, Min-Wei W, et al. (2006) Impairment of the spatial learning and memory induced by learned helplessness and chronic mild stress. Pharmacol Biochem Behav 83: 186-193. https://doi.org/10.1016/j.pbb.2006.01.004
|
| [73] |
Orsetti M, Colella L, Dellarole A, et al. (2007) Modification of spatial recognition memory and object discrimination after chronic administration of haloperidol, amitriptyline, sodium valproate or olanzapine in normal and anhedonic rats. Int J Neuropsychopharmacology 10: 345-357. https://doi.org/10.1017/S1461145706006705
|
| [74] |
Elizalde N, Gil-Bea FJ, Ramirez MJ, et al. (2008) Long-lasting behavioral effects and recognition memory deficit induced by chronic mild stress in mice: effect of antidepressant treatment. Psychopharmacology 199: 1-14. https://doi.org/10.1007/s00213-007-1035-1
|
| [75] |
Couto FS, Batalha VL, Valadas JS, et al. (2012) Escitalopram improves memory deficits induced by maternal separation in the rat. Eur J Pharmacol 695: 71-75. https://doi.org/10.1016/j.ejphar.2012.08.020
|
| [76] |
Dachir S, Kadar T, Robinzon B, et al. (1993) Cognitive deficits induced in young rats by long-term corticosterone administration. Behav Neural Biol 60: 103-109. https://doi.org/10.1016/0163-1047(93)90173-F
|
| [77] |
Coburn-Litvak PS, Pothakos K, Tata DA, et al. (2003) Chronic administration of CORT impairs spatial reference memory before spatial working memory in rats. Neurobiol Learn Mem 80: 11-23. https://doi.org/10.1016/S1074-7427(03)00019-4
|
| [78] |
Sousa N, Lukoyanov NV, Madeira MD, et al. (2000) Reorganization of the morphology of hippocampal neurites and synapses after stress-induced damage correlates with behavioral improvement. Neurosci 97: 253-266. https://doi.org/10.1016/S0306-4522(00)00050-6
|
| [79] |
van den Buuse M, Buret L, Hill R (2020) Involvement of brain-derived neurotrophic factor (BDNF) in the long-term memory effects of glucocorticoid stimulation during adolescence/young adulthood. Behav Brain Res 377: 112223. https://doi.org/10.1016/j.bbr.2019.112223
|
| [80] |
Spanagel R, Durstewitz D, Hansson A, et al. (2013) A systems medicine research approach for studying alcohol addiction. Addict Bbiol 18: 883-896. https://doi.org/10.1111/adb.12109
|
| [81] |
Frank J, Cichon S, Treutlein J, et al. (2012) Genome-wide significant association between alcohol dependence and a variant in the ADH gene cluster. Addict Biol 17: 171-180. https://doi.org/10.1111/j.1369-1600.2011.00395.x
|
| [82] |
Biernacka JM, Geske JR, Schneekloth TD, et al. (2013) Replication of genome wide association studies of alcohol dependence: support for association with variation in ADH1C. Plos One 8: e58798. https://doi.org/10.1371/journal.pone.0058798
|
| [83] |
Varlinskaya EI, Truxell EM, Spear LP (2015) Sex differences in sensitivity to the social consequences of acute ethanol and social drinking during adolescence. Behav Brain Res 282: 6-13. https://doi.org/10.1016/j.bbr.2014.12.054
|





Figures(6)

Laaziz Abderrahim, El Mostafi Hicham, Elhessni Aboubaker, Azeroil Fatima, Touil Tarik, Boumlah Soufiane, Mesfioui Abdelhalim. Sex differences in behavioral, cognitive and voluntary ethanol-intake effects in Dexamethasone-induced depression-like state in Wistar rat[J]. AIMS Neuroscience, 2022, 9(2): 228-249. doi: 10.3934/Neuroscience.2022012







 DownLoad:
DownLoad: