Transient receptor potential cation channel subfamily V member 1 (TRPV1) was considered to play pivotal roles in multiple cancers; however, the expression and clinical significance of the TRPV1 remain unclear, which were explored in this study.
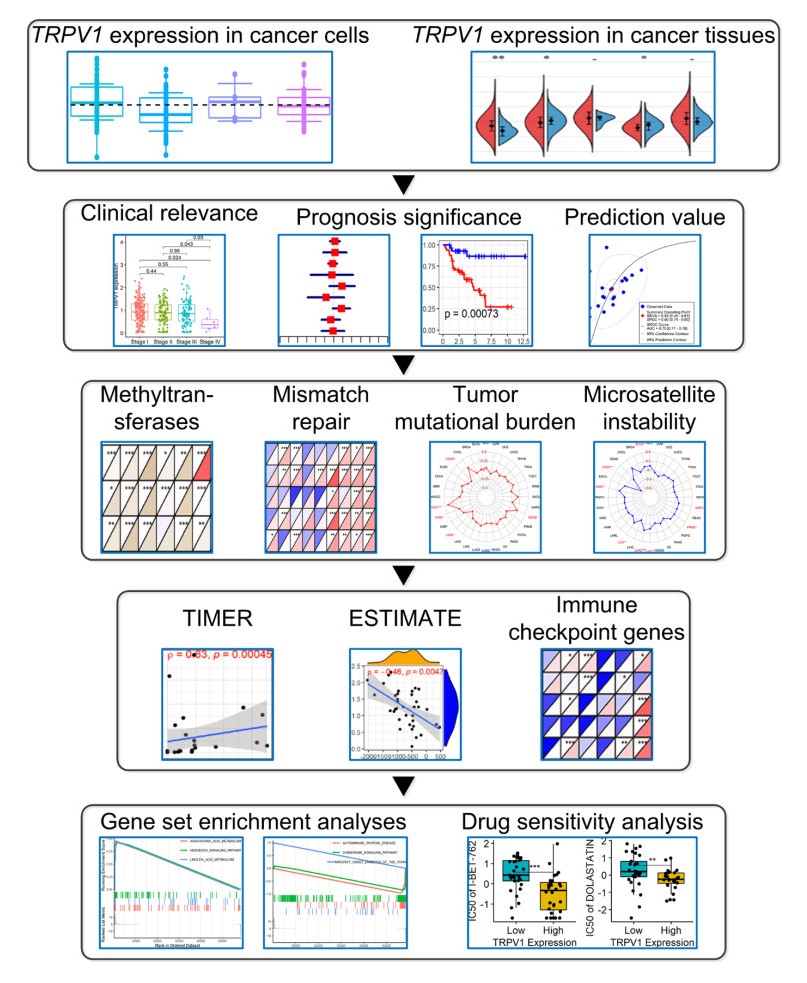
The pan-cancer analysis was performed based on 10,236 samples in 32 cancers. Differential TRPV1 expression levels were detected in 12 cancers (p < 0.05). TRPV1 demonstrated its conspicuous prognosis significance and prediction effects for some cancers (e.g., lung adenocarcinoma), indicating its potential as a valuable and novel biomarker in treating and predicting cancers. TRPV1 expression was relevant to DNA methyltransferases, mismatch repair genes, tumor mutational burden, and microsatellite instability. TRPV1 expression was associated with the immune microenvironment of some cancers, and its roles in different cancers may be mediated by affecting various immune cells. Gene set enrichment analysis discloses the significant relevance of TRPV1 expression with a series of metabolic and immunoregulatory-related pathways.
This study provided a comprehensive workflow of the expression, clinical significance, and underlying mechanisms of TRPV1 in pan-cancer. TRPV1 may be an underlying biomarker for predicting and treating multiple cancer.
Citation: Tao Huang. TRPV1 is a potential biomarker for the prediction and treatment of multiple cancers based on a pan-cancer analysis[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8361-8379. doi: 10.3934/mbe.2022389
Transient receptor potential cation channel subfamily V member 1 (TRPV1) was considered to play pivotal roles in multiple cancers; however, the expression and clinical significance of the TRPV1 remain unclear, which were explored in this study.
The pan-cancer analysis was performed based on 10,236 samples in 32 cancers. Differential TRPV1 expression levels were detected in 12 cancers (p < 0.05). TRPV1 demonstrated its conspicuous prognosis significance and prediction effects for some cancers (e.g., lung adenocarcinoma), indicating its potential as a valuable and novel biomarker in treating and predicting cancers. TRPV1 expression was relevant to DNA methyltransferases, mismatch repair genes, tumor mutational burden, and microsatellite instability. TRPV1 expression was associated with the immune microenvironment of some cancers, and its roles in different cancers may be mediated by affecting various immune cells. Gene set enrichment analysis discloses the significant relevance of TRPV1 expression with a series of metabolic and immunoregulatory-related pathways.
This study provided a comprehensive workflow of the expression, clinical significance, and underlying mechanisms of TRPV1 in pan-cancer. TRPV1 may be an underlying biomarker for predicting and treating multiple cancer.

| [1] |
F. Bray, M. Laversanne, E. Weiderpass, I. Soerjomataram, The ever-increasing importance of cancer as a leading cause of premature death worldwide, Cancer, 127 (2021), 3029-3030. https://doi.org/10.1002/cncr.33587 doi: 10.1002/cncr.33587

|
| [2] |
H. Sung, J. Ferlay, R. L. Siegel, M. Laversanne, I. Soerjomataram, A. Jemal, et al., Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA Cancer J. Clin., 71 (2021), 209-249. https://doi.org/10.3322/caac.21660 doi: 10.3322/caac.21660
|
| [3] |
W. Chen, Z. Sun, L. Lu, Targeted engineering of medicinal chemistry for cancer therapy: Recent advances and perspectives, Angew Chem. Int. Ed. Engl., 60 (2021), 5626-5643. https://doi.org/10.1002/anie.201914511 doi: 10.1002/anie.201914511
|
| [4] |
J. Berk-Krauss, J. A. Stein, J. Weber, D. Polsky, A. C. Geller, New Systematic Therapies and Trends in Cutaneous Melanoma Deaths Among US Whites, 1986-2016, Am. J. Public Health, 110 (2020), 731-733. https://doi.org/10.2105/AJPH.2020.305567 doi: 10.2105/AJPH.2020.305567
|
| [5] |
P. A. Ott, E. Elez, S. Hiret, D. W. Kim, A. Morosky, S. Saraf, et al., Pembrolizumab in patients with extensive-stage small-cell lung cancer: results from the phase Ib KEYNOTE-028 study, J. Clin. Oncol., 35 (2017), 3823-3829. https://doi.org/10.1200/JCO.2017.72.5069 doi: 10.1200/JCO.2017.72.5069
|
| [6] |
L. Li, C. Chen, C. Chiang, T. Xiao, Y. Chen, Y. Zhao, et al., The impact of TRPV1 on cancer pathogenesis and therapy: A systematic review, Int. J. Biol. Sci, . 17 (2021), 2034-2049. https://doi.org/10.7150/ijbs.59918 doi: 10.7150/ijbs.59918
|
| [7] |
K. Zhai, A. Liskova, P. Kubatka, D. Busselberg, Calcium entry through TRPV1: A potential target for the regulation of proliferation and apoptosis in cancerous and healthy cells, Int. J. Mol. Sci., 21 (2020). https://doi.org/10.3390/ijms21114177 doi: 10.3390/ijms21114177
|
| [8] |
K. Stock, J. Kumar, M. Synowitz, S. Petrosino, R. Imperatore, E. S. Smith, et al., Neural precursor cells induce cell death of high-grade astrocytomas through stimulation of TRPV1, Nat. Med., 18 (2012), 1232-1238. https://doi.org/10.1038/nm.2827 doi: 10.1038/nm.2827
|
| [9] |
L. V. Weber, K. Al-Refae, G. Wolk, G. Bonatz, J. Altmuller, C. Becker, et al., Expression and functionality of TRPV1 in breast cancer cells, Breast Cancer (Dove Med Press), 8 (2016), 243-252. https://doi.org/10.2147/BCTT.S121610 doi: 10.2147/BCTT.S121610
|
| [10] |
M. Ghandi, F. W. Huang, J. Jane-Valbuena, G. V. Kryukov, C. C. Lo, E. R. McDonald, et al., Next-generation characterization of the Cancer Cell Line Encyclopedia, Nature, 569 (2019), 503-508. https://doi.org/10.1038/s41586-019-1186-3 doi: 10.1038/s41586-019-1186-3
|
| [11] |
R. Beroukhim, C. H. Mermel, D. Porter, G. Wei, S. Raychaudhuri, J. Donovan, et al., The landscape of somatic copy-number alteration across human cancers, Nature, 463 (2010), 899-905. https://doi.org/10.1038/nature08822 doi: 10.1038/nature08822
|
| [12] |
T. Li, J. Fan, B. Wang, N. Traugh, Q. Chen, J. S. Liu, et al., TIMER: A web server for comprehensive analysis of tumor-infiltrating immune cells, Cancer Res., 77 (2017), e108-e110. https://doi.org/10.1158/0008-5472.CAN-17-0307 doi: 10.1158/0008-5472.CAN-17-0307
|
| [13] |
K. Yoshihara, M. Shahmoradgoli, E. Martinez, R. Vegesna, H. Kim, W. Torres-Garcia, et al., Inferring tumour purity and stromal and immune cell admixture from expression data, Nat. Commun., 4 (2013), 2612. https://doi.org/10.1038/ncomms3612 doi: 10.1038/ncomms3612
|
| [14] |
T. Li, J. Fu, Z. Zeng, D. Cohen, J. Li, Q. Chen, et al., TIMER2.0 for analysis of tumor-infiltrating immune cells, Nucleic Acids Res., 48 (2020), W509-W514. https://doi.org/10.1093/nar/gkaa407 doi: 10.1093/nar/gkaa407
|
| [15] |
J. Hu, B. Othmane, A. Yu, H. Li, Z. Cai, X. Chen, et al., 5mC regulator-mediated molecular subtypes depict the hallmarks of the tumor microenvironment and guide precision medicine in bladder cancer, BMC Med., 19 (2021), 289. https://doi.org/10.1186/s12916-021-02163-6 doi: 10.1186/s12916-021-02163-6
|
| [16] |
S. Fu, B. Gong, S. Wang, Q. Chen, Y. Liu, C. Zhuang, et al., Prognostic value of long noncoding rna dleu2 and its relationship with immune infiltration in kidney renal clear cell carcinoma and liver hepatocellular carcinoma, Int. J. Gen. Med., 14 (2021), 8047-8064. https://doi.org/10.2147/IJGM.S336428 doi: 10.2147/IJGM.S336428
|
| [17] |
W. Li, J. A. Ma, X. Sheng, C. Xiao Screening of CXC chemokines in the microenvironment of ovarian cancer and the biological function of CXCL10, World J. Surg. Oncol., 19 (2021), 329. https://doi.org/10.1186/s12957-021-02440-x doi: 10.1186/s12957-021-02440-x
|
| [18] |
Z. Zhuang, H. Cai, H. Lin, B. Guan, Y. Wu, Y. Zhang, et al., Development and validation of a robust pyroptosis-related signature for predicting prognosis and immune status in patients with colon cancer, J. Oncol., 2021 (2021), 5818512. https://doi.org/10.1155/2021/5818512 doi: 10.1155/2021/5818512
|
| [19] |
Y. Wang, Y. Tian, S. Liu, Z. Wang, Q. Xing, Prognostic value and immunological role of AXL gene in clear cell renal cell carcinoma associated with identifying LncRNA/RBP/AXL mRNA networks, Cancer Cell Int., 21 (2021), 625. https://doi.org/10.1186/s12935-021-02322-y doi: 10.1186/s12935-021-02322-y
|
| [20] |
Y. Zhu, Y. Zhou, H. Jiang, Z. Chen, B. Lu, Analysis of core genes for colorectal cancer prognosis based on immune and stromal scores, PeerJ, 9 (2021), e12452. https://doi.org/10.7717/peerj.12452 doi: 10.7717/peerj.12452
|
| [21] |
G. Yu, L. G. Wang, Y. Han, Q. Y. He, clusterProfiler: an R package for comparing biological themes among gene clusters, OMICS, 16 (2012), 284-287. https://doi.org/10.1089/omi.2011.0118 doi: 10.1089/omi.2011.0118
|
| [22] |
P. Langfelder, S. Horvath WGCNA: An R package for weighted correlation network analysis, BMC Bioinform., 9 (2008), 559. https://doi.org/10.1186/1471-2105-9-559 doi: 10.1186/1471-2105-9-559
|
| [23] |
W. C. Reinhold, M. Sunshine, H. Liu, S. Varma, K. W. Kohn, J. Morris, et al., CellMiner: A web-based suite of genomic and pharmacologic tools to explore transcript and drug patterns in the NCI-60 cell line set, Cancer Res., 72 (2012), 3499-3511. https://doi.org/10.1158/0008-5472.CAN-12-1370 doi: 10.1158/0008-5472.CAN-12-1370
|
| [24] |
J. Luo, L. Wu, D. Liu, Z. Xiong, L. Wang, X. Qian, et al., Gene regulatory network analysis identifies key genes and regulatory mechanisms involved in acute myocardial infarction using bulk and single cell RNA-seq data, Math. Biosci. Eng., 18 (2021), 7774-7789. https://doi.org/10.3934/mbe.2021386 doi: 10.3934/mbe.2021386
|
| [25] |
J. Cheng, J. Zhang, Z. Wu, X. Sun, Inferring microenvironmental regulation of gene expression from single-cell RNA sequencing data using scMLnet with an application to COVID-19, Brief Bioinform., 22 (2021), 988-1005. https://doi.org/10.1093/bib/bbaa327 doi: 10.1093/bib/bbaa327
|
| [26] |
G. H. Han, D. B. Chay, S. Nam, H. Cho, J. Y. Chung, J. H. Kim, Prognostic significance of Transient Receptor Potential Vanilloid Type 1 (TRPV1) and Phosphatase and Tension Homolog (PTEN) in epithelial ovarian cancer, Cancer Genom. Proteom., 17 (2020), 309-319. https://doi.org/10.21873/cgp.20191 doi: 10.21873/cgp.20191
|
| [27] |
G. H. Han, D. B. Chay, S. Nam, H. Cho, J. Y. Chung, J. H. Kim, The combination of Transient Receptor Potential Vanilloid Type 1 (TRPV1) and Phosphatase and Tension Homolog (PTEN) is an effective prognostic biomarker in cervical cancer, Int. J. Gynecol. Pathol., 40 (2021), 214-223. https://doi.org/10.1097/PGP.0000000000000677 doi: 10.1097/PGP.0000000000000677
|
| [28] |
N. Gao, F. Yang, S. Chen, H. Wan, X. Zhao, H. Dong, The role of TRPV1 ion channels in the suppression of gastric cancer development, J. Exp. Clin. Cancer Res., 39 (2020), 206. https://doi.org/10.1186/s13046-020-01707-7 doi: 10.1186/s13046-020-01707-7
|
| [29] |
M. C. Liebl, T. G. Hofmann, The Role of p53 Signaling in Colorectal Cancer, Cancers (Basel), 13 (2021). https://doi.org/10.3390/cancers13092125 doi: 10.3390/cancers13092125
|
| [30] |
V. J. N. Bykov, S. E. Eriksson, J. Bianchi, K. G. Wiman, Targeting mutant p53 for efficient cancer therapy, Nat. Rev. Cancer, 18 (2018), 89-102. https://doi.org/10.1038/nrc.2017.109 doi: 10.1038/nrc.2017.109
|
| [31] |
C. Bosson, J. Rendu, L. Pelletier, A. Abriat, A. Chatagnon, J. Brocard, et al., Variations in the TRPV1 gene are associated to exertional heat stroke, J. Sci. Med. Sport, 23 (2020), 1021-1027. https://doi.org/10.1016/j.jsams.2020.04.018 doi: 10.1016/j.jsams.2020.04.018
|
| [32] |
X. He, C. Xu, Immune checkpoint signaling and cancer immunotherapy, Cell Res., 30 (2020), 660-669. https://doi.org/10.1038/s41422-020-0343-4 doi: 10.1038/s41422-020-0343-4
|
| [33] |
M. Greally, J. F. Chou, W. K. Chatila, M. Margolis, M. Capanu, J. F. Hechtman, et al., Clinical and molecular predictors of response to immune checkpoint inhibitors in patients with advanced esophagogastric cancer, Clin. Cancer Res., 25 (2019), 6160-6169. https://doi.org/10.1158/1078-0432.CCR-18-3603 doi: 10.1158/1078-0432.CCR-18-3603
|
| [34] |
C. M. Fares, E. M. Van Allen, C. G. Drake, J. P. Allison, S. Hu-Lieskovan, Mechanisms of resistance to immune checkpoint blockade: Why does checkpoint inhibitor immunotherapy not work for all patients?, Am. Soc. Clin. Oncol. Educ. Book, 39 (2019), 147-164. https://doi.org/10.1200/EDBK_240837 doi: 10.1200/EDBK_240837
|
| [35] |
J. K. Bujak, D. Kosmala, I. M. Szopa, K. Majchrzak, P. Bednarczyk, Inflammation, Cancer and Immunity-Implication of TRPV1 Channel, Front. Oncol., 9 (2019), 1087. https://doi.org/10.3389/fonc.2019.01087 doi: 10.3389/fonc.2019.01087
|
| [36] |
S. Bertin, Y. Aoki-Nonaka, P. R. de Jong, L. L. Nohara, H. Xu, S. R. Stanwood, et al., The ion channel TRPV1 regulates the activation and proinflammatory properties of CD4(+) T cells, Nat. Immunol., 15 (2014), 1055-1063. https://doi.org/10.1038/ni.3009 doi: 10.1038/ni.3009
|
| [37] |
K. Zhang, D. Julius, Y. Cheng, Structural snapshots of TRPV1 reveal mechanism of polymodal functionality, Cell, 184 (2021), 5138-5150. https://doi.org/10.1016/j.cell.2021.08.012 doi: 10.1016/j.cell.2021.08.012
|
| [38] |
L. Pecze, B. Viskolcz, Z. Olah, Molecular surgery concept from bench to bedside: A focus on TRPV1+ pain-sensing neurons, Front. Physiol., 8 (2017), 378. https://doi.org/10.3389/fphys.2017.00378 doi: 10.3389/fphys.2017.00378
|
| [39] |
L. Pecze, W. Blum, T. Henzi, B. Schwaller, Endogenous TRPV1 stimulation leads to the activation of the inositol phospholipid pathway necessary for sustained Ca(2+) oscillations, Biochim. Biophys. Acta, 1863 (2016), 2905-2915. https://doi.org/10.1016/j.bbamcr.2016.09.013 doi: 10.1016/j.bbamcr.2016.09.013
|
| [40] |
N. Erin, Role of sensory neurons, neuroimmune pathways, and transient receptor potential vanilloid 1 (TRPV1) channels in a murine model of breast cancer metastasis, Cancer Immunol. Immunother, 69 (2020), 307-314. https://doi.org/10.1007/s00262-019-02463-0 doi: 10.1007/s00262-019-02463-0
|
| [41] |
X. Tian, W. Xu, Y. Wang, A. Anwaier, H. Wang, F. Wan, et al., Identification of tumor-infiltrating immune cells and prognostic validation of tumor-infiltrating mast cells in adrenocortical carcinoma: results from bioinformatics and real-world data, Oncoimmunology, 9 (2020), 1784529. https://doi.org/10.1080/2162402X.2020.1784529 doi: 10.1080/2162402X.2020.1784529
|
| [42] |
R. de Matos Simoes, S. Dalleau, K. E. Williamson, F. Emmert-Streib, Urothelial cancer gene regulatory networks inferred from large-scale RNAseq, Bead and Oligo gene expression data, BMC Syst. Biol., 9 (2015), 21. DOI:10.1186/s12918-015-0165-z doi: 10.1186/s12918-015-0165-z
|
 mbe-19-08-389 Supplementary.docx mbe-19-08-389 Supplementary.docx |
 |
| mbe-19-08-389 Supplementary.docx |
|







Figures(8)

Tao Huang. TRPV1 is a potential biomarker for the prediction and treatment of multiple cancers based on a pan-cancer analysis[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8361-8379. doi: 10.3934/mbe.2022389







 DownLoad:
DownLoad: