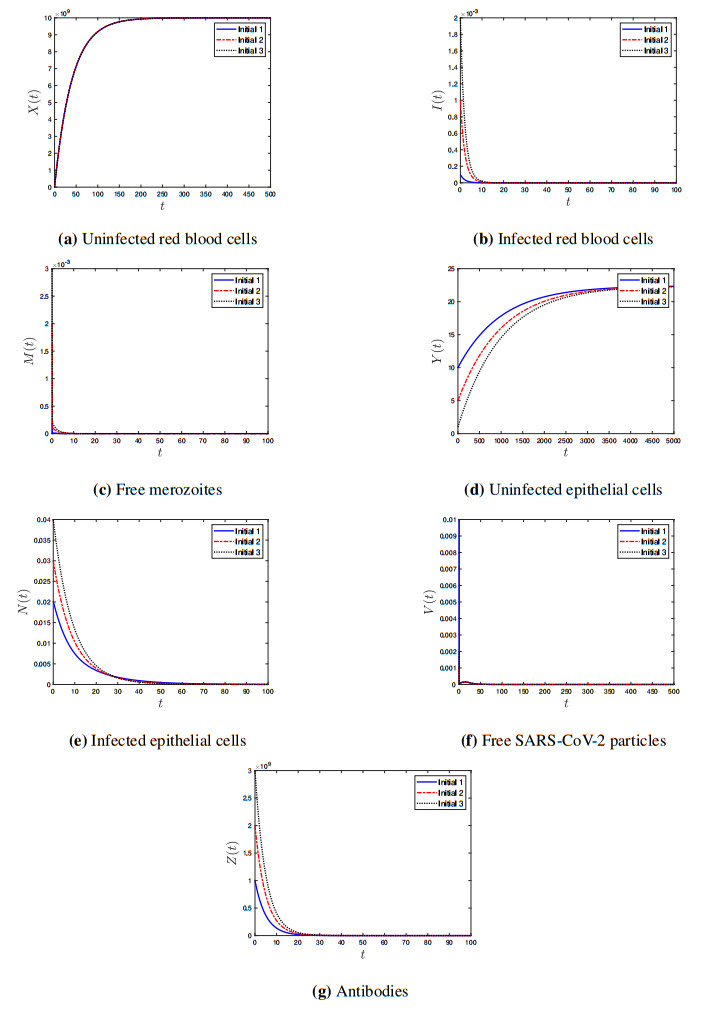
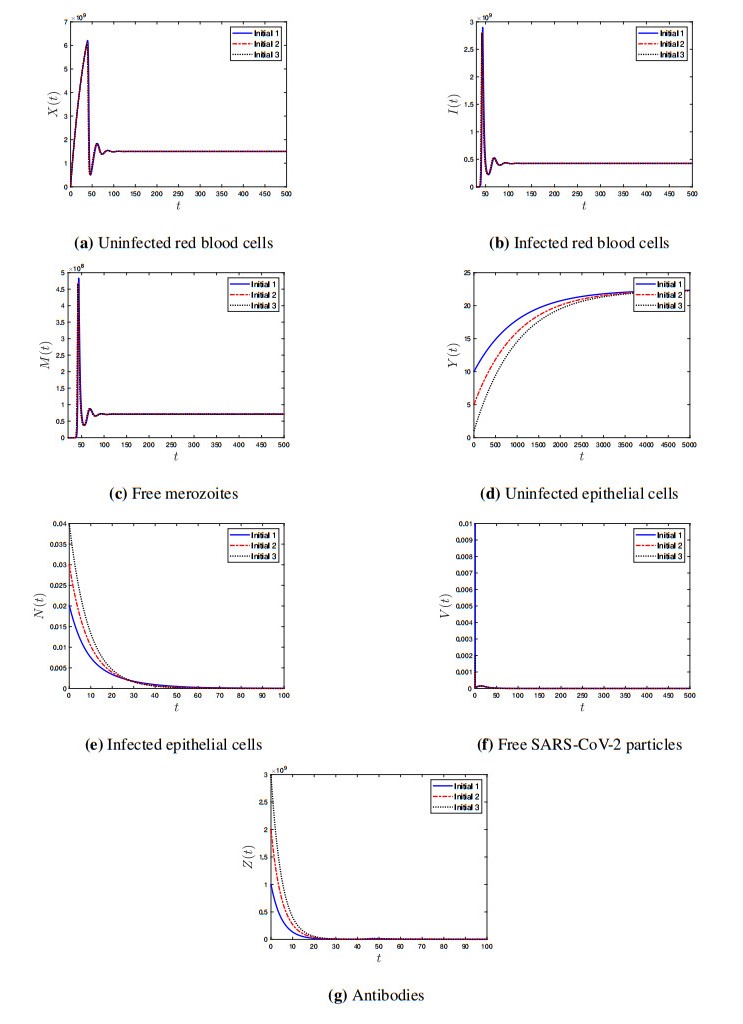
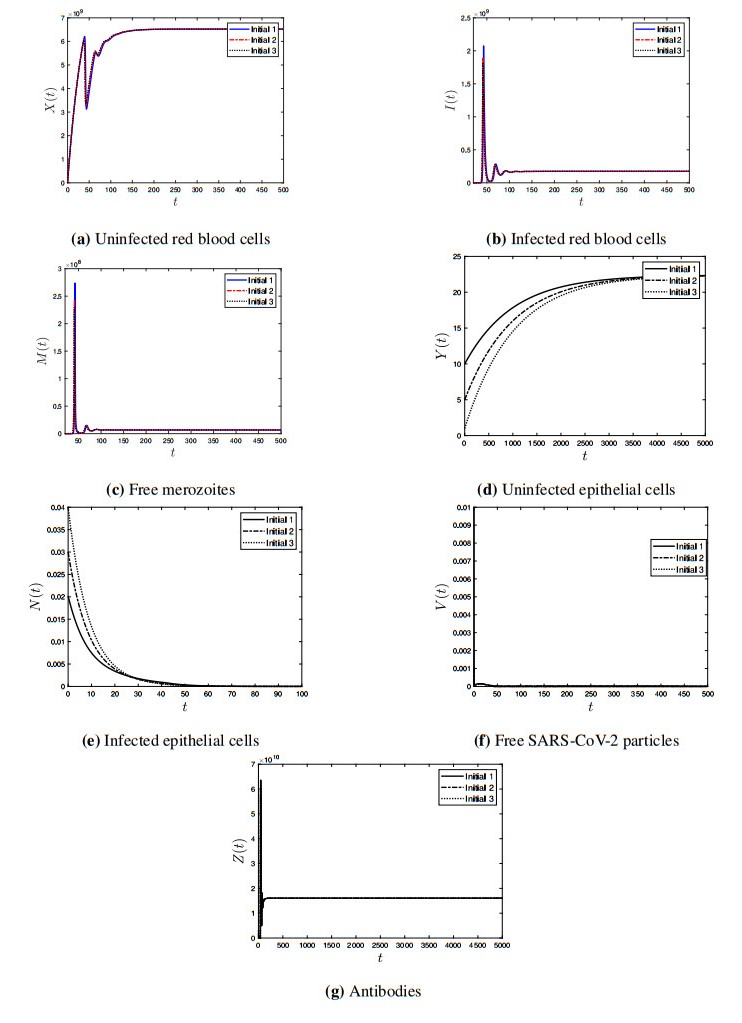
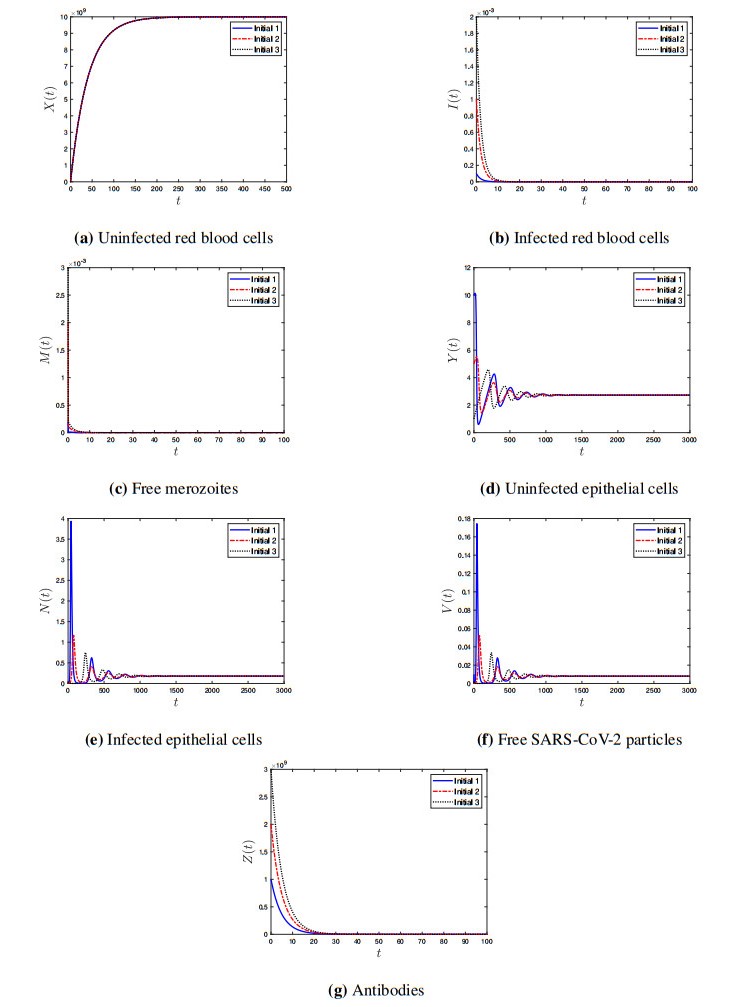
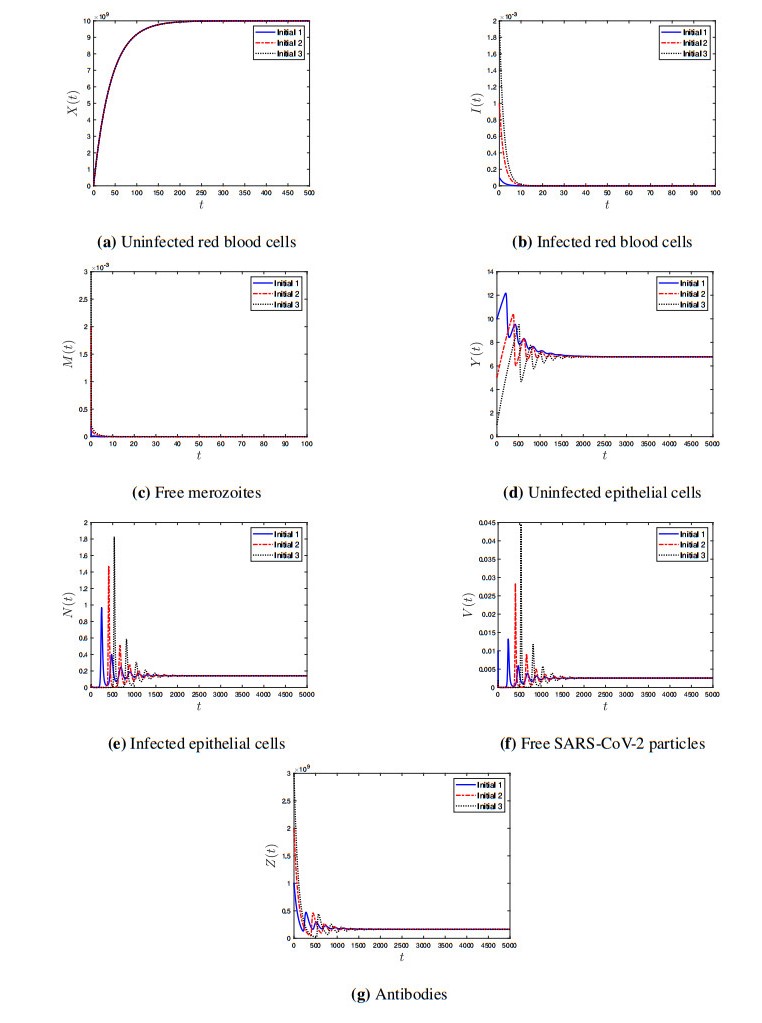
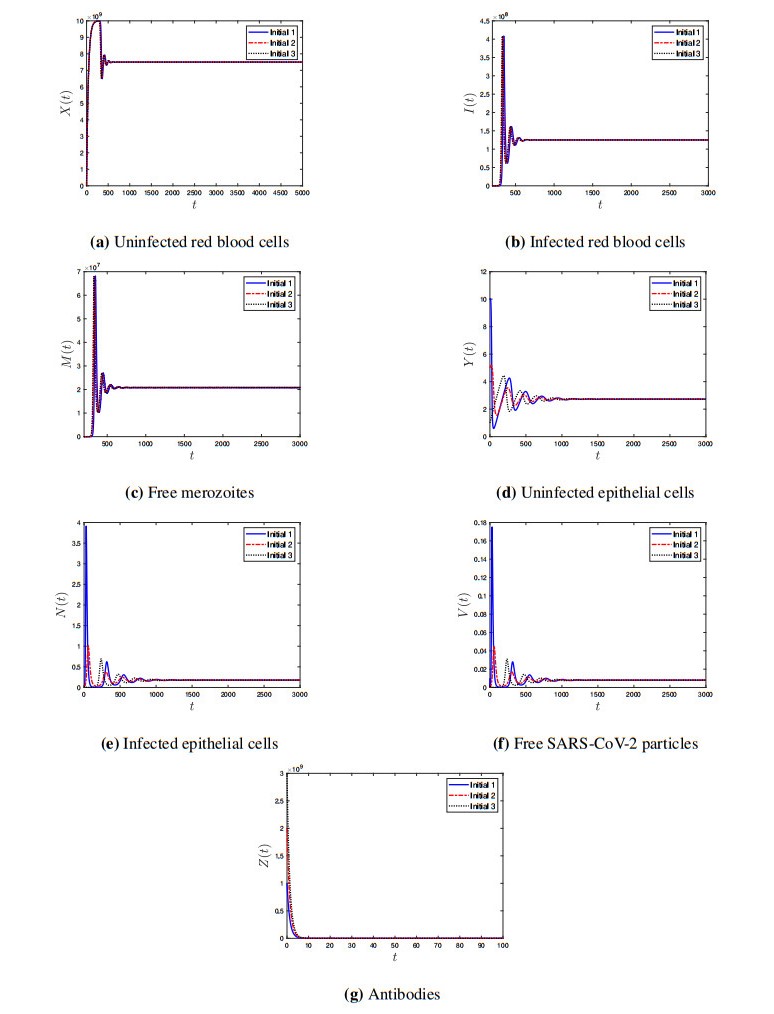
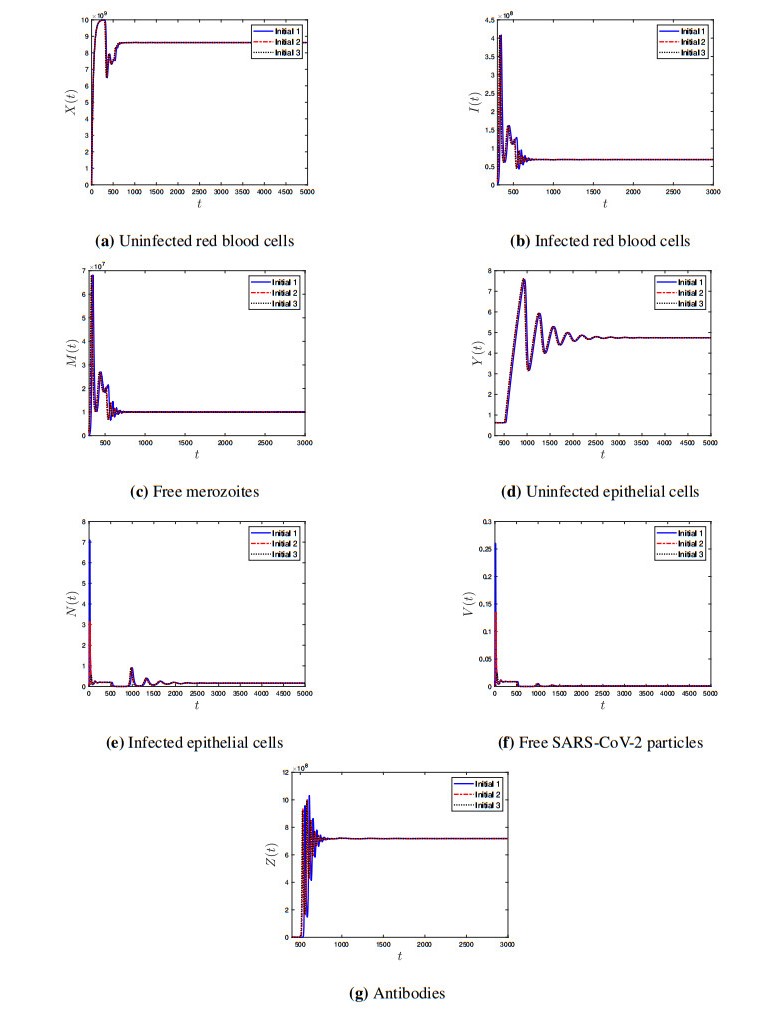
Coronavirus disease 2019 (COVID-19) is a new viral disease caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Malaria is a parasitic disease caused by Plasmodium parasites. In this paper, we explore a within-host model of SARS-CoV-2/malaria coinfection. This model consists of seven ordinary differential equations that study the interactions between uninfected red blood cells, infected red blood cells, free merozoites, uninfected epithelial cells, infected epithelial cells, free SARS-CoV-2 particles, and antibodies. We show that the model has bounded and nonnegative solutions. We compute all steady state points and derive their existence conditions. We use appropriate Lyapunov functions to confirm the global stability of all steady states. We enhance the reliability of the theoretical results by performing numerical simulations. The steady states reflect the monoinfection and coinfection with malaria and SARS-CoV-2. The shared immune response reduces the concentrations of malaria merozoites and SARS-CoV-2 particles in coinfected patients. This response reduces the severity of SARS-CoV-2 infection in this group of patients.
Citation: A. D. Al Agha, A. M. Elaiw. Global dynamics of SARS-CoV-2/malaria model with antibody immune response[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8380-8410. doi: 10.3934/mbe.2022390
Coronavirus disease 2019 (COVID-19) is a new viral disease caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Malaria is a parasitic disease caused by Plasmodium parasites. In this paper, we explore a within-host model of SARS-CoV-2/malaria coinfection. This model consists of seven ordinary differential equations that study the interactions between uninfected red blood cells, infected red blood cells, free merozoites, uninfected epithelial cells, infected epithelial cells, free SARS-CoV-2 particles, and antibodies. We show that the model has bounded and nonnegative solutions. We compute all steady state points and derive their existence conditions. We use appropriate Lyapunov functions to confirm the global stability of all steady states. We enhance the reliability of the theoretical results by performing numerical simulations. The steady states reflect the monoinfection and coinfection with malaria and SARS-CoV-2. The shared immune response reduces the concentrations of malaria merozoites and SARS-CoV-2 particles in coinfected patients. This response reduces the severity of SARS-CoV-2 infection in this group of patients.

| [1] | Coronavirus disease (COVID-19), weekly epidemiological update (5 April 2022), World Health Organization (WHO), 2022. Available from: https://www.who.int/docs/default-source/coronaviruse/situation-reports/20220405_weekly_epi_update_86.pdf?sfvrsn=3f01a460_4&download=true |
| [2] |
P. Wilairatana, F. Masangkay, K. Kotepui, G. Milanez, M. Kotepui, Prevalence and characteristics of malaria among COVID-19 individuals: A systematic review, meta-analysis, and analysis of case reports, PLoS Negl. Trop. Dis., 15 (2021), 1–18. https://doi.org/10.1371/journal.pntd.0009766 doi: 10.1371/journal.pntd.0009766

|
| [3] |
S. M. Akula, S. L. Abrams, L. S. Steelman, S. Candido, M. Librabc, K. Lerpiriyapong, et al., Cancer therapy and treatments during COVID-19 era, Adv. Biol. Regul., 77 (2020), 100739. https://doi.org/10.1016/j.jbior.2020.100739 doi: 10.1016/j.jbior.2020.100739
|
| [4] | N. Jyotsana, M. King, The impact of COVID-19 on cancer risk and treatment, Cell. Mol. Bioeng., (2020). https://doi.org/10.1155/2016/5230219 |
| [5] |
Y. Cao, The impact of the hypoxia-VEGF-vascular permeability on COVID-19-infected patients, Exploration, 1 (2021), 1–5. https://doi.org/10.1002/EXP.20210051 doi: 10.1002/EXP.20210051
|
| [6] |
M. Hussein, A. Albashir, O. Elawad, A. Homeida, Malaria and COVID-19: Unmasking their ties, Malar. J., 19 (2020), 1–10. https://doi.org/10.1186/s12936-020-03541-w doi: 10.1186/s12936-020-03541-w
|
| [7] |
F. Di Gennaro, C. Marotta, P. Locantore, D. Pizzol, G. Putoto, Malaria and COVID-19: Common and different findings, Trop. Med. Infect. Dis., 5 (2020), 1–10. https://doi.org/10.3390/tropicalmed5030141 doi: 10.3390/tropicalmed5030141
|
| [8] | Coronavirus disease (COVID-19), Vaccine tracker, World Health Organization (WHO), 2021. Available from: https://covid19.trackvaccines.org/agency/who/ |
| [9] |
A. Hussain, H. Yang, M. Zhang, Q. Liu, G. Alotaibib, M. Irfan, et al., mRNA vaccines for COVID-19 and diverse diseases, J. Control. Release, 345 (2022), 314–333. https://doi.org/10.1016/j.jconrel.2022.03.032 doi: 10.1016/j.jconrel.2022.03.032
|
| [10] | The U.S. Food and Drug Administration, Know your treatment options for COVID-19, 2021. Available from: https://www.fda.gov/consumers/consumer-updates/know-your-treatment-options-covid-19 |
| [11] |
A. M. Elaiw, A. D. Al Agha, Global analysis of a reaction-diffusion within-host malaria infection model with adaptive immune response, Mathematics, 8 (2020), 1–32. https://doi.org/10.3390/math8040563 doi: 10.3390/math8040563
|
| [12] | Malaria, Fact sheets, World Health Organization (WHO), 2021. Available from: https://www.who.int/news-room/fact-sheets/detail/malaria |
| [13] |
C. Sebastiao, C. Gaston, J. Paixao, E. Sacomboio, Z. Neto, J. N. de Vasconcelos, et al., Coinfection between SARS‐CoV‐2 and vector‐borne diseases in Luanda, Angola, J. Med. Virol., 94 (2021), 366–371. https://doi.org/10.1002/jmv.27354 doi: 10.1002/jmv.27354
|
| [14] |
A. Pusparani, J. Henrina, A. Cahyadi, Co-infection of COVID-19 and recurrent malaria, J. Infect. Dev. Ctries., 15 (2021), 625–629. https://doi.org/10.3855/jidc.13793 doi: 10.3855/jidc.13793
|
| [15] |
P. Chanda-Kapata, N. Kapata, A. Zumla, COVID-19 and malaria: A symptom screening challenge for malaria endemic countries, Int. J. Infect. Dis., 94 (2020), 151–153. https://doi.org/10.1016/j.ijid.2020.04.007 doi: 10.1016/j.ijid.2020.04.007
|
| [16] |
O. Indari, B. Baral, K. Muduli, A. Mohanty, N. Swaind, N. K. Mohakud, et al., Insights into Plasmodium and SARS-CoV-2 co-infection driven neurological manifestations, Biosafety Health, 3 (2021), 230–234. https://doi.org/10.1016/j.bsheal.2021.04.001 doi: 10.1016/j.bsheal.2021.04.001
|
| [17] |
N. Mahajan, S. Kesarwani, S. Shinde, A. Nayak, D. N. Modi, S. D. Mahale, et al., Co-infection of malaria and dengue in pregnant women with SARS-CoV-2, Int. J. Gynaecol. Obstet., 151 (2020), 459–462. https://doi.org/10.1002/ijgo.13415 doi: 10.1002/ijgo.13415
|
| [18] |
N. Mahajan, R. Gajbhiye, S. Bahirat, P. Lokhande, A. Mathe, S. Rathi, et al., Co-infection of malaria and early clearance of SARS‐CoV‐2 in healthcare workers, J. Med. Virol., 93 (2021), 2431–2438. https://doi.org/10.1002/jmv.26760 doi: 10.1002/jmv.26760
|
| [19] | R. Hussein, M. H. Guedes, N. Ibraheim, M. M. Ali, A. El-Tahir, N. Allam, et al., Impact of COVID-19 and malaria coinfection on clinical outcomes: A retrospective cohort study, Clin. Microbiol. Infect., (2022), S1198-743X(22)00167-7. https://doi.org/10.1016/j.cmi.2022.03.028 |
| [20] |
A. Kalungi, E. Kinyanda, D. Akena, P. Kaleebu, I. Bisangwa, Less severe cases of COVID-19 in Sub-Saharan Africa: Could co-infection or a recent history of Plasmodium falciparum infection be protective?, Front. Immunol., 12 (2021), 1–5. https://doi.org/10.3389/fimmu.2021.565625 doi: 10.3389/fimmu.2021.565625
|
| [21] |
A. Parodi, E. Cozzani, Coronavirus disease 2019 (COVID 19) and malaria, Med. Hypoth., 143 (2020), 110036. https://doi.org/10.1016/j.mehy.2020.110036 doi: 10.1016/j.mehy.2020.110036
|
| [22] |
M. Iesa, M. Osman, M. Hassan, A. Dirar, N. Abuzeid, J. J.Mancuso, et al., SARS-CoV-2 and Plasmodium falciparum common immunodominant regions may explain low COVID-19 incidence in the malaria-endemic belt, New Microbes New Infect., 38 (2020), 100817. https://doi.org/10.1016/j.nmni.2020.100817 doi: 10.1016/j.nmni.2020.100817
|
| [23] |
A. M. Elaiw, A. D. Al Agha, Global analysis of a reaction-diffusion within-host malaria infection model with adaptive immune response, Mathematics, 8 (2020), 1–32. https://doi.org/10.3390/math8040563 doi: 10.3390/math8040563
|
| [24] |
H. Chen, W. Wang, R. Fu, J. Luo, Global analysis of a mathematical model on malaria with competitive strains and immune responses, Appl. Math. Comput., 259 (2015), 132–152. https://doi.org/10.1016/j.amc.2015.02.073 doi: 10.1016/j.amc.2015.02.073
|
| [25] |
D. Khoury, R. Aogo, G. Randriafanomezantsoa-Radohery, J. McCaw et al., Within-host modeling of blood-stage malaria, Immunol. Rev., 285 (2018), 168–193. https://doi.org/10.1111/imr.12697 doi: 10.1111/imr.12697
|
| [26] |
F. Agusto, M. Leite, M. Orive, The transmission dynamics of a within-and between-hosts malaria model, Ecol. Complex., 38 (2019), 31–55. https://doi.org/10.1016/j.ecocom.2019.02.002 doi: 10.1016/j.ecocom.2019.02.002
|
| [27] |
T. Song, C. Wang, B. Tian, Mathematical models for within-host competition of malaria parasites, Math. Biosci. Eng., 16 (2019), 6623–6653. https://doi.org/10.3934/mbe.2019330 doi: 10.3934/mbe.2019330
|
| [28] |
E. Takoutsing, A. Temgoua, D. Yemele, S. Bowong, Dynamics of an intra-host model of malaria with periodic antimalarial treatment, Int. J. Nonlinear Sci., 27 (2019), 148–164. https://doi.org/10.11948/20160266 doi: 10.11948/20160266
|
| [29] |
A. M. Elaiw, A. D. Al Agha, Global dynamics of SARS-CoV-2/cancer model with immune responses, Appl. Math. Comput., 408 (2021), 1–19. https://doi.org/10.1016/j.amc.2021.126364 doi: 10.1016/j.amc.2021.126364
|
| [30] |
C. Li, J. Xu, J. Liu, Y. Zhou, The within-host viral kinetics of SARS-CoV-2, Math. Biosci. Eng., 17 (2020), 2853–2861. https://doi.org/10.3934/mbe.2020159 doi: 10.3934/mbe.2020159
|
| [31] | A. S. Almocera, G. Quiroz, E. A. Hernandez-Vargas, Stability analysis in COVID-19 within-host model with immune response, Commun. Nonlinear Sci. Numer. Simul., (2020), 105584. https://doi.org/10.1016/j.cnsns.2020.105584 |
| [32] |
S. Q. Du, W. Yuan, Mathematical modeling of interaction between innate and adaptive immune responses in COVID-19 and implications for viral pathogenesis, J. Med. Virol., 92 (2020), 1615–1628. https://doi.org/10.1002/jmv.25866 doi: 10.1002/jmv.25866
|
| [33] | L. Pinky, H. M. Dobrovolny, SARS-CoV-2 coinfections: Could influenza and the common cold be beneficial?, J. Med. Virol., (2020), 1–8. https://doi.org/10.1002/jmv.26098 |
| [34] |
C. Currie, J. Fowler, K. Kotiadis, T. Monks, How simulation modelling can help reduce the impact of COVID-19, J. Simul., 14 (2020), 83–97. https://doi.org/10.1080/17477778.2020.1751570 doi: 10.1080/17477778.2020.1751570
|
| [35] |
M. V. Krishna, J. Prakash, Mathematical modelling on phase based transmissibility of Coronavirus, Infect. Dis. Model., 5 (2020), 375–385. https://doi.org/10.1016/j.idm.2020.06.005 doi: 10.1016/j.idm.2020.06.005
|
| [36] |
K. Rajagopal, N. Hasanzadeh, F. Parastesh, I. Hamarash, S. Jafari, I, Hussain, A fractional-order model for the novel coronavirus (COVID-19) outbreak, Nonlinear Dyn., 101 (2020), 711–718. https://doi.org/10.1007/s11071-020-05757-6 doi: 10.1007/s11071-020-05757-6
|
| [37] |
T. Chen, J. Rui, Q. Wang, Z. Zhao, J. A. Cui, L. Yin, A mathematical model for simulating the phase-based transmissibility of a novel coronavirus, Infect. Dis. Poverty., 9 (2020), 1–8. https://doi.org/10.1186/s40249-020-00640-3 doi: 10.1186/s40249-020-00640-3
|
| [38] |
C. Yang, J. Wang, A mathematical model for the novel coronavirus epidemic in Wuhan, China, Math. Biosci. Eng., 17 (2020), 2708–2724. https://doi.org/10.3934/mbe.2020148 doi: 10.3934/mbe.2020148
|
| [39] | Z. Liu, P. Magal, O. Seydi, G. Webb, Understanding unreported cases in the 2019-nCoV epidemic outbreak in Wuhan, China, and the importance of major public health interventions, SSRN Electron. J., (2020), 1–12. http://dx.doi.org/10.2139/ssrn.3530969 |
| [40] |
N. Bellomo, R. Bingham, M. A. J. Chaplain, G. Dosi, G. Forni, D. A. Knopoff, et al., A multiscale model of virus pandemic: heterogeneous interactive entities in a globally connected world, Math. Models Methods Appl. Sci., 30 (2020), 1591–1651. https://doi.org/10.1142/S0218202520500323 doi: 10.1142/S0218202520500323
|
| [41] | H. K. Khalil, Nonlinear Systems, Prentice-Hall, New Jersey, (1996). |
| [42] |
C. Hetzel, R. Anderson, The within-host cellular dynamics of bloodstage malaria: Theoretical and experimental studies, Parasitology, 113 (1996), 25–38. https://doi.org/10.1017/s0031182000066245 doi: 10.1017/s0031182000066245
|
| [43] |
R. Anderson, R. May, S. Gupta, Non-linear phenomena in host-parasite interactions, Parasitology, 99 (1989), S59–S79. https://doi.org/10.1017/s0031182000083426 doi: 10.1017/s0031182000083426
|
| [44] |
T. Orwa, R. Mbogo, L. Luboobi, Mathematical model for the in-host malaria dynamics subject to malaria vaccines, Lett. Biomath., 5 (2018), 222–251. https://doi.org/10.1080/23737867.2018.1526132 doi: 10.1080/23737867.2018.1526132
|
| [45] |
G. T. Grassia, C. F. Markwalter, W. P. O'Meara, S. M. Taylor, A. A. Obala, SARS-CoV-2 cross-reactivity in prepandemic serum from rural malaria-infected persons, Cambodia, Emerg. Infect. Dis., 28 (2022), 1080–1081. https://doi.org/10.3201/eid2802.211725 doi: 10.3201/eid2802.211725
|
| [46] |
N. Bellomo, D. Burini, N. Outada, Multiscale models of Covid-19 with mutations and variants, Netw. Heterog. Media., 17 (2022), 293–310. https://doi.org/10.3934/nhm.2022008 doi: 10.3934/nhm.2022008
|
| [47] |
N. Bellomo, D. Burini, N. Outada, Pandemics of mutating virus and society: A multi-scale active particles approach, Philos. Trans. A. Math. Phys. Eng. Sci., 380 (2022), 1–14. https://doi.org/https://doi.org/10.1098/rsta.2021.0161 doi: 10.1098/rsta.2021.0161
|
Figures(7) / Tables(2)

A. D. Al Agha, A. M. Elaiw. Global dynamics of SARS-CoV-2/malaria model with antibody immune response[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8380-8410. doi: 10.3934/mbe.2022390







 DownLoad:
DownLoad: