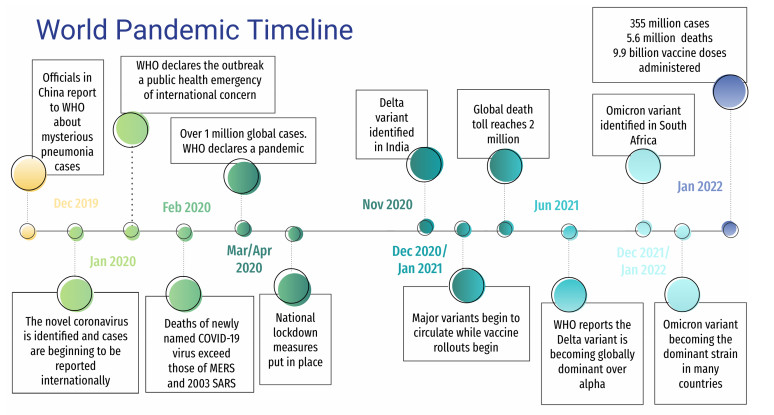
In this paper we introduce a model for the spread of COVID-19 which takes into account competing SARS-CoV-2 mutations as well as the possibility of reinfection due to fading of vaccine protection. Our primary focus is to describe the impact of the B.1.617.2 (Delta) and B.1.1.529 (Omicron) variants on the state of Hawai‘i and to illustrate how the model performed during the pandemic, both in terms of accuracy, and as a resource for the government and media. Studying the effect of the pandemic on the Hawaiian archipelago is of notable interest because, as an isolated environment, its unique geography affords partially controlled travel to and from the state. We highlight the modeling efforts of the Hawai‘i Pandemic Applied Modeling Work Group (HiPAM) which used the model presented here, and we detail the model fitting and forecasting for the periods from July 2021 to October 2021 (Delta surge) and from November 2021 to April 2022 (Omicron surge). Our results illustrate that the model was both accurate when the forecasts were built on assumptions that held true, and was inaccurate when the public response to the forecasts was to enforce safety measures that invalidated the assumptions in the model.
Citation: Richard Carney, Monique Chyba, Victoria Y. Fan, Prateek Kunwar, Thomas Lee, Ionica Macadangdang, Yuriy Mileyko. Modeling variants of the COVID-19 virus in Hawai‘i and the responses to forecasting[J]. AIMS Mathematics, 2023, 8(2): 4487-4523. doi: 10.3934/math.2023223
In this paper we introduce a model for the spread of COVID-19 which takes into account competing SARS-CoV-2 mutations as well as the possibility of reinfection due to fading of vaccine protection. Our primary focus is to describe the impact of the B.1.617.2 (Delta) and B.1.1.529 (Omicron) variants on the state of Hawai‘i and to illustrate how the model performed during the pandemic, both in terms of accuracy, and as a resource for the government and media. Studying the effect of the pandemic on the Hawaiian archipelago is of notable interest because, as an isolated environment, its unique geography affords partially controlled travel to and from the state. We highlight the modeling efforts of the Hawai‘i Pandemic Applied Modeling Work Group (HiPAM) which used the model presented here, and we detail the model fitting and forecasting for the periods from July 2021 to October 2021 (Delta surge) and from November 2021 to April 2022 (Omicron surge). Our results illustrate that the model was both accurate when the forecasts were built on assumptions that held true, and was inaccurate when the public response to the forecasts was to enforce safety measures that invalidated the assumptions in the model.

| [1] | American Hospital Association: CDC estimates 58% of Americans have been infected with SARS-CoV-2, Apr. 26, 2022. Available from: https://www.aha.org/news/headline/2022-04-26-cdc-estimates-58-americans-have-been-infected-sars-cov-2 |
| [2] |
S. Allred, M. Chyba, J. M. Hyman, Y. Mileyko, B. Piccoli, The COVID-19 Pandemic Evolution in Hawai'i and New Jersey: A Lesson on Infection Transmissibility and the Role of Human Behavior, Predicting Pandemics in a Globally Connected World, 1 (2022), 109–140. https://doi.org/10.1007/978-3-030-96562-4_4 doi: 10.1007/978-3-030-96562-4_4

|
| [3] | N. Bellomo, M. A. J. Chaplain, Predicting Pandemics in a Globally Connected World, Volume 1 Toward a Multiscale, Multidisciplinary Framework through Modeling and Simulation, Springer International Publishing, 2022. https://doi.org/10.1007/978-3-030-96562-4 |
| [4] | Bloomberg: Omicron Four Times More Transmissible Than Delta in New Study, By Kanoko Matsuyama, Dec. 8, 2021. Available from: https://www.bloomberg.com/news/articles/2021-12-09/omicron-four-times-more-transmissible-than-delta-in-japan-study |
| [5] | C. Branas, A. Rundle, S. Pei, W. Yang, B. G. Carr, S. Sims, et al., Flattening the curve before it flattens us: hospital critical care capacity limits and mortality from novel coronavirus (SARS-CoV2) cases in US counties, medRxiv, 2020. |
| [6] |
D. Buitrago-Garcia, D. Egli-Gany, M. J. Counotte, S. Hossmann, H. Imeri, A. M. Ipekci, et al., Occurrence and transmission potential of asymptomatic and presymptomatic SARS-CoV-2 infections: A living systematic review and meta-analysis, PLOS Med., 17 (2020), e1003346. https://doi.org/10.1371/journal.pmed.1003346 doi: 10.1371/journal.pmed.1003346
|
| [7] | M. Chyba, Y. Mileyko, O. Markovichenko, R. Carney, A. Koniges, Epidemiological Model of the Spread of COVID-19 in Hawaii's Challenging Fight Against the Disease, The Ninth International Conference on Global Health Challenges, (2020), 32–38. |
| [8] | M. Chyba, T. Klotz, Y. Mileyko, C. Shanbrom, A look at endemic equilibria of compartmental epidemiological models and model control via vaccination and mitigation, arXiv: 2109.01738, 2021. |
| [9] |
M. Chyba, P. Kunwar, Y. Mileyko, A. Tong, W. Lau, A. Koniges, COVID-19 heterogeneity in islands chain environment, PLoS One, 17 (2022), e0263866. https://doi.org/10.1371/journal.pone.0263866 doi: 10.1371/journal.pone.0263866
|
| [10] | N. M. Ferguson, D. Laydon, G. Nedjati-Gilani, N. Imai, K. Ainslie, M. Baguelin, et al., Report 9 - Impact of non-pharmaceutical interventions (NPIs) to reduce COVID-19 mortality and healthcare demand, Imperial College London, 10 (2020), 491–197. |
| [11] |
M. Gatto, E. Bertuzzo, L. Mari, S. Miccoli, L. Carraro, R. Casagrandi, et al., Spread and dynamics of the COVID-19 epidemic in Italy: Effects of emergency containment measures, Proceedings of the National Academy of Sciences, 117 (2020), 10484–10491. https://doi.org/10.1073/pnas.2004978117 doi: 10.1073/pnas.2004978117
|
| [12] |
G. Giordano, F. Blanchini, R. Bruno, P. Colaneri, A. Di Filippo, A. Di Matteo, et al., Modelling the COVID-19 epidemic and implementation of population-wide interventions in Italy, Nat. Med., 26 (2020), 855–860. https://doi.org/10.1038/s41591-020-0883-7 doi: 10.1038/s41591-020-0883-7
|
| [13] |
W. T. Harvey, A. M. Carabelli, B. Jackson, R. K. Gupta, E. C. Thomson, E. M. Harrison, et al., SARS-CoV-2 variants, spike mutations and immune escape, Nat. Rev. Microbiol., 19 (2021), 409–424. https://doi.org/10.1038/s41579-021-00573-0 doi: 10.1038/s41579-021-00573-0
|
| [14] | Hawaii Department of Health, Disease Outbreak Control Division: COVID-19 Dashboard. Available from: https://health.hawaii.gov/coronavirusdisease2019 |
| [15] | Hawaii Population Model. Hawai'i Data Collaborative, Department of Health, Disease Outbreak Control Division: COVID-19 Data Reports, 2022. Available from: https://health.hawaii.gov/coronavirusdisease2019/what-you-should-know/covid-19-data-reports |
| [16] | Hawai'i Department of Health: Vaccination Distribution Plan, 2022. Available from: www.hawaiicovid19.com/wp-content/uploads/2021/01/Executive-Summary_Final1_010721.pdf |
| [17] | Hawaii News Now: Ventilators from Hawaii's emergency stockpile now in use as more relief nurses are on their way, By Allyson Blair, Aug. 25, 2021. Available from: https://www.hawaiinewsnow.com/2021/08/26/ventilators-emergency-stockpile-now-use-morerelief-nurses-are-their-way/ |
| [18] | Hawai'i Pandemic Applied Modeling Work Group, 2022. Available from: www.hipam.org |
| [19] |
H. W. Hethcote, The Mathematics of Infectious Diseases, SIAM Rev., 42 (2000), 599–653. https://doi.org/10.1137/S0036144500371907 doi: 10.1137/S0036144500371907
|
| [20] | Honolulu Star-Advertiser: Hawaii's daily COVID cases are forecast to peak at 3,700 in October, By Nina Wu, Aug. 23, 2021. Available from: https://www.staradvertiser.com/2021/08/23/hawaiinews/hawaiis-daily-covid-cases-are-forecast-to-peak-at-3700-in-october/ |
| [21] | Honolulu Star-Advertiser: Kaiser postpones elective surgeries and procedures on Oahu, Maui due to COVID surge, By Nina Wu, Aug. 27, 2021. Available from: https://www.staradvertiser.com/2021/08/27/breaking-news/kaiser-postpones-elective-surgeriesand-procedures-on-oahu-maui-due-to-covid-surge/ |
| [22] | Honolulu Star-Advertiser: No ICU beds available at Queen's medical facilities as COVID cases surge in Hawaii, By Sophie Cocke, Aug. 17, 2021. Available from: https://www.staradvertiser.com/2021/08/17/hawaii-news/no-icu-beds-available-at-queensmedical-facilities-as-covid-cases-surge-in-hawaii/ |
| [23] |
A. D. Iuliano, J. M. Brunkard, T. K. Boehmer, E. Peterson, S. Adjei, A. M. Binder, et al., Trends in Disease Severity and Health Care Utilization During the Early Omicron Variant Period Compared with Previous SARS-CoV-2 High Transmission Periods — United States, December 2020–January 2022, MMWR Morb. Mortal., 71 (2022), 146–152. https://doi.org/10.15585/mmwr.mm7104e4 doi: 10.15585/mmwr.mm7104e4
|
| [24] | V. Kala, K. Guo, E. Swantek, A. Tong, M. Chyba, Y. Mileyko, et al., Pandemics in Hawai‘i: 1918 Influenza and COVID-19, The Ninth International Conference on Global Health Challenges, (2020), 26–31. |
| [25] | D. G. Kendall, Deterministic and stochastic epidemics in closed populations, University of California Press, (2020), 149–166. https://doi.org/10.1525/9780520350717-011 |
| [26] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, R. Soc. Lond. A, 115 (1927), 700–721. https://doi.org/10.1098/rspa.1927.0118 doi: 10.1098/rspa.1927.0118
|
| [27] |
W. O. Kermack, A. G. McKendrick, Contributions to the mathematical theory of epidemics. Ⅱ.—The problem of endemicity, Proceedings of the Royal Society of London. Series A, containing papers of a mathematical and physical character, 138 (1932), 55–83. https://doi.org/10.1098/rspa.1932.0171 doi: 10.1098/rspa.1932.0171
|
| [28] |
P. Kunwar, O. Markovichenko, M. Chyba, Y. Mileyko, A. Koniges, T. Lee, A study of computational and conceptual complexities of compartment and agent based models, Netw. Heterog. Media, 17 (2022), 359–384. https://doi.org/10.3934/nhm.2022011 doi: 10.3934/nhm.2022011
|
| [29] |
J. O. Lloyd-Smith, A. P. Galvani, W. M. Getz, Curtailing transmission of severe acute respiratory syndrome within a community and its hospital, Proceedings of the Royal Society of London. Series B: Biological Sciences, 270 (2003), 1979–1989. https://doi.org/10.1098/rspb.2003.2481 doi: 10.1098/rspb.2003.2481
|
| [30] |
N. Ortega, M. Ribes, M. Vidal, R. Rubio, R. Aguilar, S. Williams, et al, Seven-month kinetics of SARS-CoV-2 antibodies and role of pre-existing antibodies to human coronaviruses, Nat. Commun., 12 (2021), 1–10. https://doi.org/10.1038/s41467-021-24979-9 doi: 10.1038/s41467-021-24979-9
|
| [31] |
M. Park, A. R. Cook, J. T. Lim, Y. Sun, B. L. Dickens, A Systematic Review of COVID-19 Epidemiology Based on Current Evidence, J. Clin. Med., 9 (2020), 967. https://doi.org/10.3390/jcm9040967 doi: 10.3390/jcm9040967
|
| [32] |
P. Qu, J. N. Faraone, J. P. Evans, Y. M. Zheng, L. Yu, Q. Ma, et al., Durability of Booster mRNA Vaccine against SARS- CoV-2 BA.2.12.1, BA.4, and BA.5 Subvariants, New Engl. J. Med., 387 (2022), 1329–1331. https://doi.org/10.1056/NEJMc2210546 doi: 10.1056/NEJMc2210546
|
| [33] | A. Richterman, E. A. Meyerowitz, M. Cevik, Indirect Protection by Reducing Transmission: Ending the Pandemic With Severe Acute Respiratory Syndrome Coronavirus 2 Vaccination, Open Forum Infectious Diseases Open Forum Infectious Diseases, 9(2) (2022), ofab259. https://doi.org/10.1093/ofid/ofab259 |
| [34] |
R. Ross, H. P. Hudson, An Application of the Theory of Probabilities to the Study of a priori Pathometry. Part Ⅱ, Proceedings of the Royal Society of London Series A, 93 (1917), 212–225. https://doi.org/10.1098/rspa.1917.0014 doi: 10.1098/rspa.1917.0014
|
| [35] |
R. Ross, An Application of the Theory of Probabilities to the Study of a priori Pathometry. Part Ⅰ, Proceedings of the Royal Society of London Series A, 92 (1916), 204–230. https://doi.org/10.1098/rspa.1916.0007 doi: 10.1098/rspa.1916.0007
|
| [36] |
Y. B. Ruhomally, M. Mungur, A. A. H. Khoodaruth, V. Oree, M. Z. Dauhoo, Assessing the Impact of Contact Tracing, Quarantine and Red Zone on the Dynamical Evolution of the Covid-19 Pandemic using the Cellular Automata Approach and the Resulting Mean Field System: A Case study in Mauritius, Appl. Math. Model., 111 (2022), 567–589. https://doi.org/10.1016/j.apm.2022.07.008 doi: 10.1016/j.apm.2022.07.008
|
| [37] | Science, Science Insider, Health: More people are getting COVID-19 twice, suggesting immunity wanes quickly in some, By Jop De Vrieze, Nov. 18, 2020. Available from: https://www.science.org/content/article/more-people-are-getting-covid-19-twice-suggestingimmunity-wanes-quickly-some |
| [38] | M. T. Sofonea, B. Roquebert, V. Foulongne, L. Verdurme, S. Trombert-Paolantoni, M. Roussel, et al., From Delta to Omicron: analysing the SARS-CoV-2 epidemic in France using variant-specific screening tests (September 1 to December 18, 2021) Cold Spring Harbor Laboratory Press, 2022. https://www.medrxiv.org/content/10.1101/2021.12.31.21268583v1 |
| [39] |
S. Sturniolo, W. Waites, T. Colbourn, D. Manheim, J. Panovska-Griffiths, Testing, tracing and isolation in compartmental models, PLoS Comput. Biol., 17 (2021), e1008633. https://doi.org/10.1371/journal.pcbi.1008633 doi: 10.1371/journal.pcbi.1008633
|
| [40] | State of Hawai'i Portal: Travel Data. Available from: https://hawaiicovid19.com/travel/data/ |
| [41] | S. T. Tan, A. T. Kwan, I. Rodríguez-Barraquer, B. J. Singer, H. J. Park, J. A. Lewnard, et al., Infectiousness of SARS-CoV-2 breakthrough infections and reinfections during the Omicron wave, medRxiv 2022.08.08.22278547. |
| [42] | U.S. Bureau of Labor Statistics: Healthcare Population, May 2020, State Occupational Employment and Wage Estimates Hawaii. Available from: https://www.bls.gov/oes/current/oeshi.htm29-0000 |
| [43] | U.S. Census Bureau. Available from: https://data.census.gov/cedsci/ |
| [44] |
F. Zhou, T. Yu, R. Du, G. Fan, Y. Liu, Z. Liu, et al., Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study, The Lancet, 395 (2020), 1054–1062. https://doi.org/10.1016/S0140-6736(20)30566-3 doi: 10.1016/S0140-6736(20)30566-3
|























Figures(24) / Tables(10)

Richard Carney, Monique Chyba, Victoria Y. Fan, Prateek Kunwar, Thomas Lee, Ionica Macadangdang, Yuriy Mileyko. Modeling variants of the COVID-19 virus in Hawai‘i and the responses to forecasting[J]. AIMS Mathematics, 2023, 8(2): 4487-4523. doi: 10.3934/math.2023223







 DownLoad:
DownLoad: