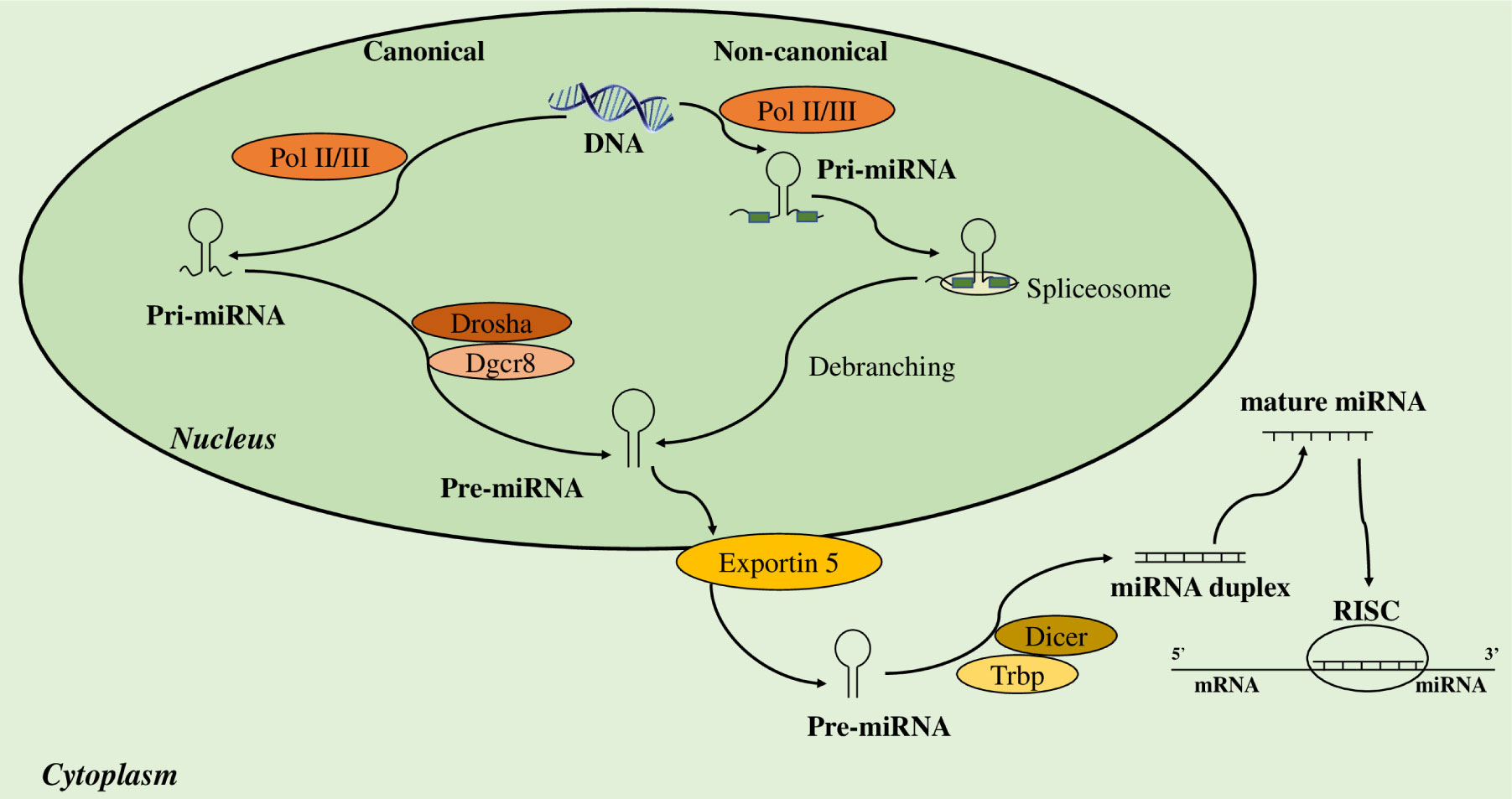
Wound healing (WH) is a fundamental physiological process to keep the integrity of the skin, therefore impaired and chronic WH is a common and severe medical problem and represent one of the biggest challenges of public health. The resolution of the WH inflammatory phase is characterized by a complex series of events that involves many cellular types, especially neutrophils, macrophages and inflammatory mediators, which are crucial for a correct wound closure. MicroRNAs (miRNAs) play essential roles in wound repair. In fact, miR-142 is linked to inflammation modulating neutrophils' chemotaxis and polarization, while the polarization of M1 toward the M2 phenotype is driven by miR-223 and miR-132 is linked to chemokines and cytokines that activate endothelial cells and attract leukocytes and peripheral cells to the damage site. Thus, understanding the dysregulation of miRNAs in WH will be decisive for the development of new and more effective therapies for the management of chronic wounds.
Citation: Federica Serra, Lisa Aielli, Erica Costantini. The role of miRNAs in the inflammatory phase of skin wound healing[J]. AIMS Allergy and Immunology, 2021, 5(4): 264-278. doi: 10.3934/Allergy.2021020
Wound healing (WH) is a fundamental physiological process to keep the integrity of the skin, therefore impaired and chronic WH is a common and severe medical problem and represent one of the biggest challenges of public health. The resolution of the WH inflammatory phase is characterized by a complex series of events that involves many cellular types, especially neutrophils, macrophages and inflammatory mediators, which are crucial for a correct wound closure. MicroRNAs (miRNAs) play essential roles in wound repair. In fact, miR-142 is linked to inflammation modulating neutrophils' chemotaxis and polarization, while the polarization of M1 toward the M2 phenotype is driven by miR-223 and miR-132 is linked to chemokines and cytokines that activate endothelial cells and attract leukocytes and peripheral cells to the damage site. Thus, understanding the dysregulation of miRNAs in WH will be decisive for the development of new and more effective therapies for the management of chronic wounds.

| [1] |
Liu J, Shu B, Zhou Z, et al. (2020) Involvement of miRNA203 in the proliferation of epidermal stem cells during the process of DM chronic wound healing through Wnt signal pathways. Stem Cell Res Ther 11: 348. doi: 10.1186/s13287-020-01829-x

|
| [2] |
Li D, Peng H, Qu L, et al. (2021) miR-19a/b and miR-20a promote wound healing by regulating the inflammatory response of keratinocytes. J Invest Dermatol 141: 659-671. doi: 10.1016/j.jid.2020.06.037
|
| [3] |
Xu J, Wu W, Zhang L, et al. (2012) The role of microRNA-146a in the pathogenesis of the diabetic wound-healing impairment: correction with mesenchymal stem cell treatment. Diabetes 61: 2906-2912. doi: 10.2337/db12-0145
|
| [4] |
Wu J, Li X, Li D, et al. (2020) MicroRNA-34 family enhances wound inflammation by targeting LGR4. J Invest Dermatol 140: 465-476. doi: 10.1016/j.jid.2019.07.694
|
| [5] |
O'Brien J, Hayder H, Zayed Y, et al. (2018) Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol 9: 402. doi: 10.3389/fendo.2018.00402
|
| [6] |
Lu TX, Rothenberg ME (2018) MicroRNA. J Allergy Clin Immun 141: 1202-1207. doi: 10.1016/j.jaci.2017.08.034
|
| [7] |
Ramalingam P, Palanichamy JK, Singh A, et al. (2014) Biogenesis of intronic miRNAs located in clusters by independent transcription and alternative splicing. RNA 20: 76-87. doi: 10.1261/rna.041814.113
|
| [8] |
Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281-297. doi: 10.1016/S0092-8674(04)00045-5
|
| [9] |
Vishnoi A, Rani S (2017) MiRNA Biogenesis and regulation of diseases: An overview. Methods Mol Biol 1509: 1-10. doi: 10.1007/978-1-4939-6524-3_1
|
| [10] |
Johanson TM, Lew AM, Chong MM (2013) MicroRNA-independent roles of the RNase III enzymes Drosha and Dicer. Open Biol 3: 130144. doi: 10.1098/rsob.130144
|
| [11] |
Zhang L, Lu Q, Chang C (2020) Epigenetics in health and disease. Adv Exp Med Biol 1253: 3-55. doi: 10.1007/978-981-15-3449-2_1
|
| [12] |
Saliminejad K, Khorram Khorshid HR, Soleymani Fard S, et al. (2019) An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J Cell Physiol 234: 5451-5465. doi: 10.1002/jcp.27486
|
| [13] |
Krol J, Loedige I, Filipowicz W (2010) The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet 11: 597-610. doi: 10.1038/nrg2843
|
| [14] |
Lao TD, Le TAH (2020) MicroRNAs: Biogenesis, functions and potential biomarkers for early screening, prognosis and therapeutic molecular monitoring of nasopharyngeal carcinoma. Processes 8: 966. doi: 10.3390/pr8080966
|
| [15] |
Xu J, Chen Q, Zen K, et al. (2013) Synaptosomes secrete and uptake functionally active microRNAs via exocytosis and endocytosis pathways. J Neurochem 124: 15-25. doi: 10.1111/jnc.12057
|
| [16] |
Ganju A, Khan S, Hafeez BB, et al. (2017) miRNA nanotherapeutics for cancer. Drug Discov Today 22: 424-432. doi: 10.1016/j.drudis.2016.10.014
|
| [17] |
Rousselle P, Braye F, Dayan G (2019) Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv Drug Deliver Rev 146: 344-365. doi: 10.1016/j.addr.2018.06.019
|
| [18] |
Wang Y, Feng Z, Yang M, et al. (2021) Discovery of a novel short peptide with efficacy in accelerating the healing of skin wounds. Pharmacol Res 163: 105296. doi: 10.1016/j.phrs.2020.105296
|
| [19] |
Dini V, Janowska A, Oranges T, et al. (2020) Surrounding skin management in venous leg ulcers: A systematic review. J Tissue Viability 29: 169-175. doi: 10.1016/j.jtv.2020.02.004
|
| [20] |
Lindley LE, Stojadinovic O, Pastar I, et al. (2016) Biology and biomarkers for wound healing. Plast Reconstr Surg 138: 18S-28S. doi: 10.1097/PRS.0000000000002682
|
| [21] |
Gosain A, DiPietro LA (2004) Aging and wound healing. World J Surg 28: 321-326. doi: 10.1007/s00268-003-7397-6
|
| [22] |
Han G, Ceilley R (2017) Chronic wound healing: A review of current management and treatments. Adv Ther 34: 599-610. doi: 10.1007/s12325-017-0478-y
|
| [23] |
Etulain J (2018) Platelets in wound healing and regenerative medicine. Platelets 29: 556-568. doi: 10.1080/09537104.2018.1430357
|
| [24] |
Rodrigues M, Kosaric N, Bonham CA, et al. (2019) Wound healing: A cellular perspective. Physiol Rev 99: 665-706. doi: 10.1152/physrev.00067.2017
|
| [25] |
Opneja A, Kapoor S, Stavrou EX (2019) Contribution of platelets, the coagulation and fibrinolytic systems to cutaneous wound healing. Thromb Res 179: 56-63. doi: 10.1016/j.thromres.2019.05.001
|
| [26] |
Reinke JM, Sorg H (2012) Wound repair and regeneration. Eur Surg Res 49: 35-43. doi: 10.1159/000339613
|
| [27] |
Wilkinson HN, Hardman MJ (2020) Wound healing: cellular mechanisms and pathological outcomes. Open Biol 10: 200223. doi: 10.1098/rsob.200223
|
| [28] |
Velnar T, Bailey T, Smrkolj V (2009) The wound healing process: An overview of the cellular and molecular mechanisms. J Int Med Res 37: 1528-1542. doi: 10.1177/147323000903700531
|
| [29] |
de Oliveira S, Rosowski EE, Huttenlocher A (2016) Neutrophil migration in infection and wound repair: going forward in reverse. Nat Rev Immunol 16: 378-391. doi: 10.1038/nri.2016.49
|
| [30] |
Gonzalez AC, Costa TF, Andrade ZA, et al. (2016) Wound healing—A literature review. An Bras Dermatol 91: 614-620. doi: 10.1590/abd1806-4841.20164741
|
| [31] |
Thuraisingam T, Xu YZ, Eadie K, et al. (2010) MAPKAPK-2 signaling is critical for cutaneous wound healing. J Invest Dermatol 130: 278-286. doi: 10.1038/jid.2009.209
|
| [32] |
Werner S, Krieg T, Smola H (2007) Keratinocyte-fibroblast interactions in wound healing. J Invest Dermatol 127: 998-1008. doi: 10.1038/sj.jid.5700786
|
| [33] |
Jevtić M, Löwa A, Nováčková A, et al. (2020) Impact of intercellular crosstalk between epidermal keratinocytes and dermal fibroblasts on skin homeostasis. Biochim Biophys Acta Mol Cell Res 1867: 118722. doi: 10.1016/j.bbamcr.2020.118722
|
| [34] |
Broughton G, Janis JE, Attinger CE (2006) The basic science of wound healing. Plast Reconstr Surg 117: 12S-34S. doi: 10.1097/01.prs.0000225430.42531.c2
|
| [35] |
Demidova-Rice TN, Hamblin MR, Herman IM (2012) Acute and impaired wound healing: pathophysiology and current methods for drug delivery, part 1: normal and chronic wounds: biology, causes, and approaches to care. Adv Skin Wound Care 25: 304-314. doi: 10.1097/01.ASW.0000416006.55218.d0
|
| [36] |
Lang H, Zhao F, Zhang T, et al. (2017) MicroRNA-149 contributes to scarless wound healing by attenuating inflammatory response. Mol Med Rep 16: 2156-2162. doi: 10.3892/mmr.2017.6796
|
| [37] |
Raziyeva K, Kim Y, Zharkinbekov Z, et al. (2021) Immunology of acute and chronic wound healing. Biomolecules 11: 700. doi: 10.3390/biom11050700
|
| [38] |
Fordham JB, Naqvi AR, Nares S (2015) Regulation of miR-24, miR-30b, and miR-142-3p during macrophage and dendritic cell differentiation potentiates innate immunity. J Leukocyte Biol 98: 195-207. doi: 10.1189/jlb.1A1014-519RR
|
| [39] |
Tahamtan A, Teymoori-Rad M, Nakstad B, et al. (2018) Anti-inflammatory MicroRNAs and their potential for inflammatory diseases treatment. Front Immunol 9: 1377. doi: 10.3389/fimmu.2018.01377
|
| [40] |
Mori R, Tanaka K, Shimokawa I (2018) Identification and functional analysis of inflammation-related miRNAs in skin wound repair. Dev Growth Differ 60: 306-315. doi: 10.1111/dgd.12542
|
| [41] |
Tanaka K, Kim SE, Yano H, et al. (2017) MiR-142 is required for staphylococcus aureus clearance at skin wound sites via small GTPase-mediated regulation of the neutrophil actin cytoskeleton. J Invest Dermatol 137: 931-940. doi: 10.1016/j.jid.2016.11.018
|
| [42] | de Kerckhove M, Tanaka K, Umehara T, et al. (2018) Targeting miR-223 in neutrophils enhances the clearance of Staphylococcus aureus in infected wounds. EMBO Mol Med 10: e9024. |
| [43] |
Li D, Wang A, Liu X, et al. (2015) MicroRNA-132 enhances transition from inflammation to proliferation during wound healing. J Clin Invest 125: 3008-3026. doi: 10.1172/JCI79052
|
| [44] |
Xu X, Hong P, Wang Z, et al. (2021) MicroRNAs in transforming growth factor-beta signaling pathway associated with fibrosis involving different systems of the human body. Front Mol Biosci 8: 707461. doi: 10.3389/fmolb.2021.707461
|
| [45] |
Li D, Landén NX (2017) MicroRNAs in skin wound healing. Eur J Dermatol 27: 12-14. doi: 10.1684/ejd.2017.3040
|
| [46] |
Mahesh G, Biswas R (2019) MicroRNA-155: A master regulator of inflammation. J Interferon Cytokine Res 39: 321-330. doi: 10.1089/jir.2018.0155
|
| [47] |
Cao M, Shikama Y, Kimura H, et al. (2017) Mechanisms of impaired neutrophil migration by MicroRNAs in myelodysplastic syndromes. J Immunol 198: 1887-1899. doi: 10.4049/jimmunol.1600622
|
| [48] |
Wang CR, Zhu HF, Zhu Y (2019) Knockout of MicroRNA-155 ameliorates the Th17/Th9 immune response and promotes wound healing. Curr Med Sci 39: 954-964. doi: 10.1007/s11596-019-2128-x
|
| [49] |
Yang LL, Liu JQ, Bai XZ, et al. (2014) Acute downregulation of miR-155 at wound sites leads to a reduced fibrosis through attenuating inflammatory response. Biochem Biophys Res Commun 453: 153-159. doi: 10.1016/j.bbrc.2014.09.077
|
| [50] |
Ye J, Kang Y, Sun X, et al. (2017) MicroRNA-155 inhibition promoted wound healing in diabetic rats. Int J Low Extrem Wounds 16: 74-84. doi: 10.1177/1534734617706636
|
| [51] | Eissa MG, Artlett CM (2019) The MicroRNA miR-155 is essential in fibrosis. Noncoding RNA 5: 23. |
| [52] |
Fujiwara N, Kobayashi K (2005) Macrophages in inflammation. Curr Drug Targets Inflammation Allergy 4: 281-286. doi: 10.2174/1568010054022024
|
| [53] |
Dang CP, Leelahavanichkul A (2020) Over-expression of miR-223 induces M2 macrophage through glycolysis alteration and attenuates LPS-induced sepsis mouse model, the cell-based therapy in sepsis. PLoS One 15: e0236038. doi: 10.1371/journal.pone.0236038
|
| [54] |
Meng ZX, Wang GX, Lin JD (2012) A microRNA circuitry links macrophage polarization to metabolic homeostasis. Circulation 125: 2815-2917. doi: 10.1161/CIRCULATIONAHA.112.111518
|
| [55] |
Das A, Ganesh K, Khanna S, et al. (2014) Engulfment of apoptotic cells by macrophages: a role of MicroRNA-21 in the resolution of wound inflammation. J Immunol 192: 1120-1129. doi: 10.4049/jimmunol.1300613
|
| [56] |
Sheedy FJ (2015) Turning 21: induction of miR-21 as a key switch in the inflammatory response. Front Immunol 6: 19. doi: 10.3389/fimmu.2015.00019
|
| [57] |
Banerjee J, Sen CK (2013) MicroRNAs in skin and wound healing. Methods Mol Biol 936: 343-356. doi: 10.1007/978-1-62703-083-0_26
|
| [58] |
Lai WF, Siu PM (2014) MicroRNAs as regulators of cutaneous wound healing. J Biosciences 39: 519-524. doi: 10.1007/s12038-014-9421-4
|
| [59] |
Liechty C, Hu J, Zhang L, et al. (2020) Role of microRNA-21 and its underlying mechanisms in inflammatory responses in diabetic wounds. Int J Mol Sci 21: 3328. doi: 10.3390/ijms21093328
|
| [60] |
Wang JK, Wang Z, Li G (2019) MicroRNA-125 in immunity and cancer. Cancer Lett 454: 134-145. doi: 10.1016/j.canlet.2019.04.015
|
| [61] |
Tili E, Michaille JJ, Cimino A, et al. (2007) Modulation of miR-155 and miR-125b levels following lipopolysaccharide/TNF-alpha stimulation and their possible roles in regulating the response to endotoxin shock. J Immunol 179: 5082-5089. doi: 10.4049/jimmunol.179.8.5082
|
| [62] |
Meng Z, Zhou D, Gao Y, et al. (2018) miRNA delivery for skin wound healing. Adv Drug Deliver Rev 129: 308-318. doi: 10.1016/j.addr.2017.12.011
|
| [63] |
Hoppstädter J, Dembek A, Linnenberger R, et al. (2019) Toll-Like receptor 2 release by macrophages: An anti-inflammatory program induced by glucocorticoids and lipopolysaccharide. Front Immunol 10: 1634. doi: 10.3389/fimmu.2019.01634
|
| [64] |
Lee HM, Kim TS, Jo EK (2016) MiR-146 and miR-125 in the regulation of innate immunity and inflammation. BMB Rep 49: 311-318. doi: 10.5483/BMBRep.2016.49.6.056
|
| [65] |
Oliveira-Nascimento L, Massari P, Wetzler LM (2012) The role of TLR2 in infection and immunity. Front Immunol 3: 79. doi: 10.3389/fimmu.2012.00079
|
| [66] |
Meisgen F, Landén NX, Bouez C, et al. (2014) Activation of toll-like receptors alters the microRNA expression profile of keratinocytes. Exp Dermatol 23: 281-283. doi: 10.1111/exd.12361
|
| [67] |
Zhang X, Liu J, Wu L, et al. (2020) MicroRNAs of the miR-17~92 family maintain adipose tissue macrophage homeostasis by sustaining IL-10 expression. Elife 9: e55676. doi: 10.7554/eLife.55676
|
| [68] |
Li D, Peng H, Qu L, et al. (2021) miR-19a/b and miR-20a promote wound healing by regulating the inflammatory response of keratinocytes. J Invest Dermatol 141: 659-671. doi: 10.1016/j.jid.2020.06.037
|
| [69] |
Zhang C, Qian D, Zhao H, et al. (2018) MiR17 improves insulin sensitivity through inhibiting expression of ASK1 and anti-inflammation of macrophages. Biomed Pharmacother 100: 448-454. doi: 10.1016/j.biopha.2018.02.012
|
| [70] | Li H, Han X, Zuo K, et al. (2018) miR-23b promotes cutaneous wound healing through inhibition of the inflammatory responses by targeting ASK1. Acta Biochim Biophys Sin 50: 1104-1113. |
| [71] |
Zhao F, Lang H, Wang Z, et al. (2019) Human novel MicroRNA Seq-915_x4024 in keratinocytes contributes to skin regeneration by suppressing scar formation. Mol Ther Nucleic Acids 14: 410-423. doi: 10.1016/j.omtn.2018.12.016
|
| [72] |
Ban E, Jeong S, Park M, et al. (2020) Accelerated wound healing in diabetic mice by miRNA-497 and its anti-inflammatory activity. Biomed Pharmacother 121: 109613. doi: 10.1016/j.biopha.2019.109613
|
| [73] |
Bajan S, Hutvagner G (2020) RNA-based therapeutics: From antisense oligonucleotides to miRNAs. Cells 9: 137. doi: 10.3390/cells9010137
|
| [74] |
Li X, Li D, Wang A, et al. (2017) MicroRNA-132 with therapeutic potential in chronic wounds. J Invest Dermatol 137: 2630-2638. doi: 10.1016/j.jid.2017.08.003
|
| [75] |
Devalliere J, Chang WG, Andrejecsk JW, et al. (2014) Sustained delivery of proangiogenic microRNA-132 by nanoparticle transfection improves endothelial cell transplantation. FASEB J 28: 908-922. doi: 10.1096/fj.13-238527
|
| [76] |
Simonson B, Das S (2015) MicroRNA therapeutics: the next magic bullet? Mini Rev Med Chem 15: 467-474. doi: 10.2174/1389557515666150324123208
|


Figures(3)

Federica Serra, Lisa Aielli, Erica Costantini. The role of miRNAs in the inflammatory phase of skin wound healing[J]. AIMS Allergy and Immunology, 2021, 5(4): 264-278. doi: 10.3934/Allergy.2021020







 DownLoad:
DownLoad: