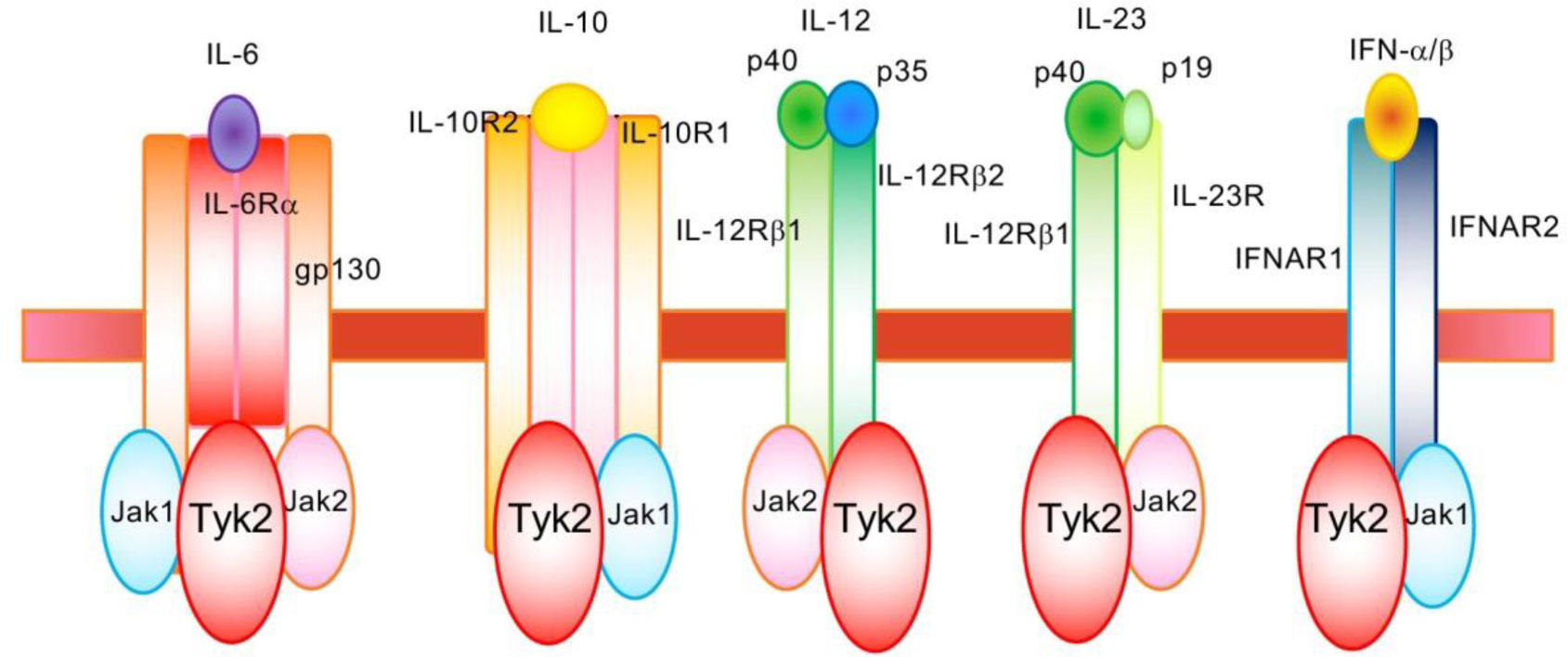
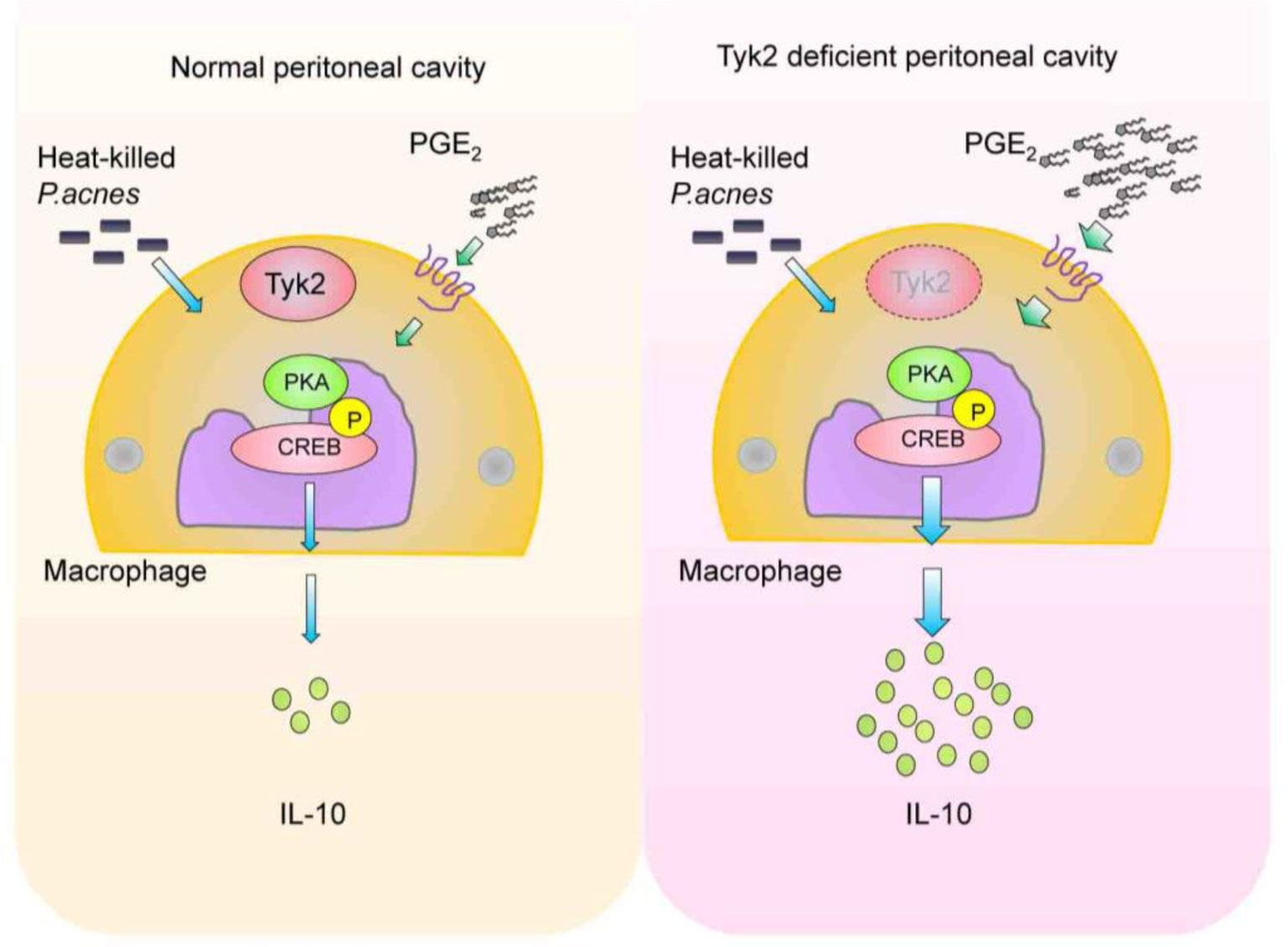
Tyrosine kinase 2 (Tyk2), which associates with the receptors for type I interferon (IFN) and interleukins (IL)-6, IL-10, IL-12, and IL-23, is critical to mediate cytokine-induced signals. Tyk2 plays an essential role in the constitutive production of small amount of type I IFNs and in the promotion of differentiation from naïve T cells into Th1 or Th17 effector cells via IL-12- and IL-23-induced signals. Additionally, Tyk2-mediated signaling suppresses the in vivo production of IL-10, which is a strong anti-inflammatory cytokine. The elevated IL-10 production in the peritoneal cells of Tyk2-deficient mice are alleviated by treatment with either diclofenac, a cyclooxygenase inhibitor, or H-89, a protein kinase A inhibitor. Notably, significantly higher basal prostaglandin E2 (PGE2) production is observed in peritoneal cavity of Tyk2-deficient mice than that of wild-type mice. Phosphorylation of cAMP response element-binding protein, induced by P. acnes and PGE2 addition, is upregulated in Tyk2-deficient macrophages. This indicates that higher IL-10 production in Tyk2-deficient mice is likely a result of the enhanced PGE2-protein kinase A pathway. Thus, Tyk2-mediated signaling regulates multiple events during immune and/or inflammatory responses.
Citation: Ryuta Muromoto, Kenji Oritani, Tadashi Matsuda. Tyk2-mediated homeostatic control by regulating the PGE2-PKA-IL-10 axis[J]. AIMS Allergy and Immunology, 2021, 5(3): 175-183. doi: 10.3934/Allergy.2021013
Tyrosine kinase 2 (Tyk2), which associates with the receptors for type I interferon (IFN) and interleukins (IL)-6, IL-10, IL-12, and IL-23, is critical to mediate cytokine-induced signals. Tyk2 plays an essential role in the constitutive production of small amount of type I IFNs and in the promotion of differentiation from naïve T cells into Th1 or Th17 effector cells via IL-12- and IL-23-induced signals. Additionally, Tyk2-mediated signaling suppresses the in vivo production of IL-10, which is a strong anti-inflammatory cytokine. The elevated IL-10 production in the peritoneal cells of Tyk2-deficient mice are alleviated by treatment with either diclofenac, a cyclooxygenase inhibitor, or H-89, a protein kinase A inhibitor. Notably, significantly higher basal prostaglandin E2 (PGE2) production is observed in peritoneal cavity of Tyk2-deficient mice than that of wild-type mice. Phosphorylation of cAMP response element-binding protein, induced by P. acnes and PGE2 addition, is upregulated in Tyk2-deficient macrophages. This indicates that higher IL-10 production in Tyk2-deficient mice is likely a result of the enhanced PGE2-protein kinase A pathway. Thus, Tyk2-mediated signaling regulates multiple events during immune and/or inflammatory responses.

| [1] |
O'Shea JJ, Ma A, Lipsky P (2002) Cytokines and autoimmunity. Nat Rev Immunol 2: 37-45. doi: 10.1038/nri702

|
| [2] |
Stark GR, Darnell JE (2012) The JAK-STAT pathway at twenty. Immunity 36: 503-514. doi: 10.1016/j.immuni.2012.03.013
|
| [3] |
Velazquez L, Fellous M, Stark GR, et al. (1992) A protein tyrosine kinase in the interferon alpha/beta signaling pathway. Cell 70: 313-322. doi: 10.1016/0092-8674(92)90105-L
|
| [4] |
Shimoda K, Kato K, Aoki K, et al. (2000) Tyk2 plays a restricted role in IFN alpha signaling, although it is required for IL-12-mediated T cell function. Immunity 13: 561-571. doi: 10.1016/S1074-7613(00)00055-8
|
| [5] |
Karaghiosoff M, Neubauer H, Lassnig C, et al. (2000) Partial impairment of cytokine responses in Tyk2-deficient mice. Immunity 3: 549-560. doi: 10.1016/S1074-7613(00)00054-6
|
| [6] |
Ishizaki M, Akimoto T, Muromoto R, et al. (2011) Involvement of tyrosine kinase-2 in both the IL-12/Th1 and IL-23/Th17 axes in vivo. J Immunol 187: 181-189. doi: 10.4049/jimmunol.1003244
|
| [7] |
Schwartz DM, Kanno Y, Villarino A, et al. (2017) JAK inhibition as a therapeutic strategy for immune and inflammatory diseases. Nat Rev Drug Discov 16: 843-862. doi: 10.1038/nrd.2017.201
|
| [8] |
Karaghiosoff M, Steinborn R, Kovarik P, et al. (2003) Central role for type I interferons and Tyk2 in lipopolysaccharide-induced endotoxin shock. Nat Immunol 4: 471-477. doi: 10.1038/ni910
|
| [9] |
Kamezaki K, Shimoda K, Numata A, et al. (2004) The role of Tyk2, Stat1 and Stat4 in LPS-induced endotoxin signals. Int Immunol 16: 1173-117. doi: 10.1093/intimm/dxh118
|
| [10] |
Vogl C, Flatt T, Fuhrmann B, et al. (2010) Transcriptome analysis reveals a major impact of JAK protein tyrosine kinase 2 (Tyk2) on the expression of interferon-responsive and metabolic genes. BMC Genomics 11: 199. doi: 10.1186/1471-2164-11-199
|
| [11] |
Shimoda K, Kamesaki K, Numata A, et al. (2002) Cutting edge: tyk2 is required for the induction and nuclear translocation of Daxx which regulates IFN-alpha-induced suppression of B lymphocyte formation. J Immunol 169: 4707-4711. doi: 10.4049/jimmunol.169.9.4707
|
| [12] |
Ishizaki M, Muromoto R, Akimoto T, et al. (2011) Tyk2 deficiency protects joints against destruction in anti-type II collagen antibody-induced arthritis in mice. Int Immunol 23: 575-558. doi: 10.1093/intimm/dxr057
|
| [13] |
Minegishi Y, Saito M, Morio T, et al. (2006) Human tyrosine kinase 2 deficiency reveals its requisite roles in multiple cytokine signals involved in innate and acquired immunity. Immunity 25: 745-755. doi: 10.1016/j.immuni.2006.09.009
|
| [14] |
Seto Y, Nakajima H, Suto A, et al. (2003) Enhanced Th2 cell-mediated allergic inflammation in Tyk2-deficient mice. J Immunol 170: 1077-1083. doi: 10.4049/jimmunol.170.2.1077
|
| [15] |
Murphy KM, Reiner SL (2002) The lineage decisions of helper T cells. Nat Rev Immunol 2: 933-944. doi: 10.1038/nri954
|
| [16] |
Korn T, Bettelli E, Oukka M, et al. (2009) IL-17 and Th17 Cells. Annu Rev Immunol 27: 485-517. doi: 10.1146/annurev.immunol.021908.132710
|
| [17] |
Ueno A, Jeffery L, Kobayashi T, et al. (2018) Th17 plasticity and its relevance to inflammatory bowel disease. J Autoimmun 87: 38-49. doi: 10.1016/j.jaut.2017.12.004
|
| [18] |
Yang P, Qian FY, Zhang MF, et al. (2019) Th17 cell pathogenicity and plasticity in rheumatoid arthritis. J Leukocyte Biol 106: 1233-1240. doi: 10.1002/JLB.4RU0619-197R
|
| [19] |
Oyamada A, Ikebe H, Itsumi M, et al. (2009) Tyrosine kinase 2 plays critical roles in the pathogenic CD4 T cell responses for the development of experimental autoimmune encephalomyelitis. J Immunol 183: 7539-7546. doi: 10.4049/jimmunol.0902740
|
| [20] |
Tokumasa N, Suto A, Kagami S, et al. (2007) Expression of Tyk2 in dendritic cells is required for IL-12, IL-23, and IFN-gamma production and the induction of Th1 cell differentiation. Blood 110: 553-560. doi: 10.1182/blood-2006-11-059246
|
| [21] |
Ishizaki M, Muromoto R, Akimoto T, et al. (2014) Tyk2 is a therapeutic target for psoriasis-like skin inflammation. Int Immunol 26: 257-267. doi: 10.1093/intimm/dxt062
|
| [22] |
Gough DJ, Messina NL, Clarke CJ, et al. (2012) Constitutive type I interferon modulates homeostatic balance through tonic signaling. Immunity 36: 166-174. doi: 10.1016/j.immuni.2012.01.011
|
| [23] |
Taniguchi T, Takaoka A (2001) A weak signal for strong responses: interferon-alpha/beta revisited. Nat Rev Mol Cell Bio 2: 378-386. doi: 10.1038/35073080
|
| [24] |
Fleetwood AJ, Dinh H, Cook AD, et al. (2009) GM-CSF- and M-CSF-dependent macrophage phenotypes display differential dependence on type I interferon signaling. J Leukocyte Biol 86: 411-421. doi: 10.1189/jlb.1108702
|
| [25] |
Tanaka T, Yamamoto Y, Muromoto R, et al. (2011) PDLIM2 inhibits T helper 17 cell development and granulomatous inflammation through degradation of STAT3. Sci Signal 4: ra85. doi: 10.1126/scisignal.2001637
|
| [26] |
Hirashima K, Muromoto R, Minoguchi H, et al. (2020) The mechanism of Tyk2 deficiency-induced immunosuppression in mice involves robust IL-10 production in macrophages. Cytokine 130: 155077. doi: 10.1016/j.cyto.2020.155077
|
| [27] |
Chang EY, Guo B, Doyle SE, et al. (2007) Cutting edge: involvement of the type I IFN production and signaling pathway in lipopolysaccharide-induced IL-10 production. J Immunol 178: 6705-6709. doi: 10.4049/jimmunol.178.11.6705
|
| [28] |
Izumi K, Mine K, Inoue Y, et al. (2015) Reduced Tyk2 gene expression in β-cells due to natural mutation determines susceptibility to virus-induced diabetes. Nat Commun 6: 6748. doi: 10.1038/ncomms7748
|
| [29] |
Nagafuchi S, Kamada-Hibio Y, Hirakawa K, et al. (2015) TYK2 Promoter Variant and Diabetes Mellitus in the Japanese. EBioMedicine 2: 744-749. doi: 10.1016/j.ebiom.2015.05.004
|
| [30] |
Mine K, Hirakawa K, Kondo S, et al. (2017) Subtyping of type 1 diabetes as classified by anti-GAD antibody, IgE levels, and tyrosine kinase 2 (TYK2) promoter variant in the Japanese. EBioMedicine 23: 46-51. doi: 10.1016/j.ebiom.2017.08.012
|
| [31] |
Papp K, Gordon K, Thaçi D, et al. (2018) Phase 2 Trial of Selective Tyrosine Kinase 2 Inhibition in Psoriasis. N Engl J Med 379: 1313-1321. doi: 10.1056/NEJMoa1806382
|

Figures(3)

Ryuta Muromoto, Kenji Oritani, Tadashi Matsuda. Tyk2-mediated homeostatic control by regulating the PGE2-PKA-IL-10 axis[J]. AIMS Allergy and Immunology, 2021, 5(3): 175-183. doi: 10.3934/Allergy.2021013







 DownLoad:
DownLoad: