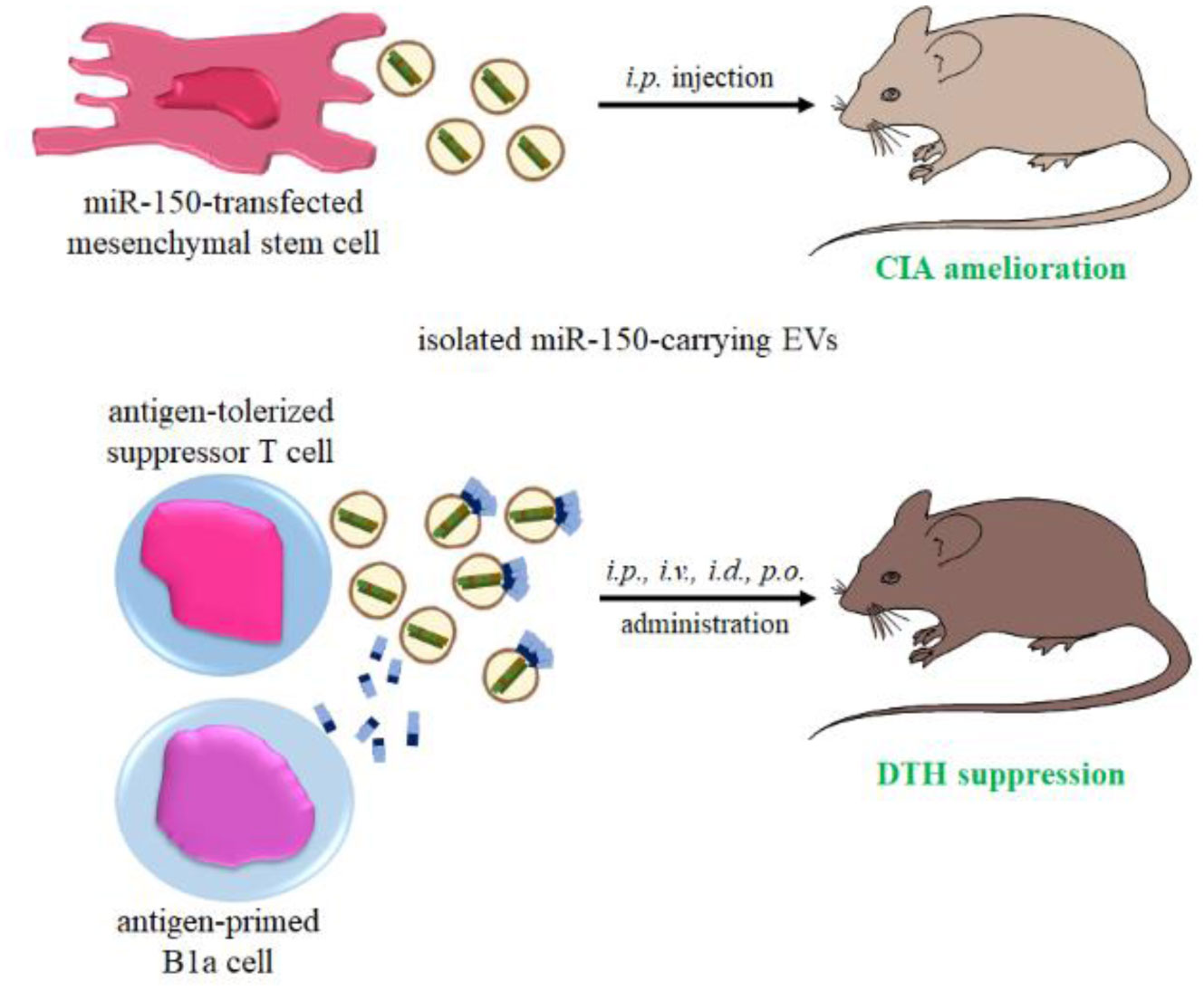
At present, special efforts are being made to develop the strategies allowing for activation of long-lasting antigen-specific immune tolerance in therapy of allergic and autoimmune diseases. Some of these therapeutic approaches are aimed at modulating cell functions at genetic level by using miRNA-based and miRNA-targeting treatments. Simultaneously, the crucial role of extracellular vesicles as natural miRNA conveyors is highlighted for induction of antigen-specific immune tolerance, especially that they appear to be easily manipulatable for therapeutic applications. Among other immune-related miRNAs, miR-150 is getting special attention as it is differently expressed by immune cells at various stages of their maturation and differentiation. In addition, miR-150 is involved in different signaling cascades orchestrating humoral and cell-mediated mechanisms of both innate and adaptive immune responses. Therefore, miR-150 is considered a master regulator of immunity in mammals. Currently, physiological miR-150-dependent regulatory circuits and causes of their malfunctioning that underlie the pathogenesis of allergic and autoimmune disorders are being unraveled. Thus, present review summarizes the current knowledge of the role of miR-150 in the pathogenesis and complications of these diseases. Furthermore, the involvement of miR-150 in regulation of immune responses to allergens and self-antigens and in induction of antigen-specific immune tolerance is discussed with the special emphasis on the therapeutic potential of this miRNA.
Citation: Katarzyna Nazimek. The complex functions of microRNA-150 in allergy, autoimmunity and immune tolerance[J]. AIMS Allergy and Immunology, 2021, 5(4): 195-221. doi: 10.3934/Allergy.2021016
At present, special efforts are being made to develop the strategies allowing for activation of long-lasting antigen-specific immune tolerance in therapy of allergic and autoimmune diseases. Some of these therapeutic approaches are aimed at modulating cell functions at genetic level by using miRNA-based and miRNA-targeting treatments. Simultaneously, the crucial role of extracellular vesicles as natural miRNA conveyors is highlighted for induction of antigen-specific immune tolerance, especially that they appear to be easily manipulatable for therapeutic applications. Among other immune-related miRNAs, miR-150 is getting special attention as it is differently expressed by immune cells at various stages of their maturation and differentiation. In addition, miR-150 is involved in different signaling cascades orchestrating humoral and cell-mediated mechanisms of both innate and adaptive immune responses. Therefore, miR-150 is considered a master regulator of immunity in mammals. Currently, physiological miR-150-dependent regulatory circuits and causes of their malfunctioning that underlie the pathogenesis of allergic and autoimmune disorders are being unraveled. Thus, present review summarizes the current knowledge of the role of miR-150 in the pathogenesis and complications of these diseases. Furthermore, the involvement of miR-150 in regulation of immune responses to allergens and self-antigens and in induction of antigen-specific immune tolerance is discussed with the special emphasis on the therapeutic potential of this miRNA.

| [1] |
O'Brien J, Hayder H, Zayed Y, et al. (2018) Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol 9: 402. doi: 10.3389/fendo.2018.00402

|
| [2] |
Cardinal-Fernández P, Ferruelo A, Esteban A, et al. (2016) Characteristics of microRNAs and their potential relevance for the diagnosis and therapy of the acute respiratory distress syndrome: from bench to bedside. Transl Res 169: 102-111. doi: 10.1016/j.trsl.2015.11.004
|
| [3] |
Dexheimer PJ, Cochella L (2020) MicroRNAs: from mechanism to organism. Front Cell Dev Biol 8: 409. doi: 10.3389/fcell.2020.00409
|
| [4] |
Nazimek K, Bryniarski K (2020) Approaches to inducing antigen-specific immune tolerance in allergy and autoimmunity: Focus on antigen-presenting cells and extracellular vesicles. Scand J Immunol 91: e12881. doi: 10.1111/sji.12881
|
| [5] |
Nazimek K, Bryniarski K (2020) Perspectives in manipulating EVs for therapeutic applications: focus on cancer treatment. Int J Mol Sci 21: 4623. doi: 10.3390/ijms21134623
|
| [6] |
Monticelli S, Ansel KM, Xiao C, et al. (2005) MicroRNA profiling of the murine hematopoietic system. Genome Biol 6: R71. doi: 10.1186/gb-2005-6-8-r71
|
| [7] |
Zhou B, Wang S, Mayr C, et al. (2007) miR-150, a microRNA expressed in mature B and T cells, blocks early B cell development when expressed prematurely. P Natl Acad Sci USA 104: 7080-7085. doi: 10.1073/pnas.0702409104
|
| [8] |
Xiao C, Calado DP, Galler G, et al. (2007) MiR-150 controls B cell differentiation by targeting the transcription factor c-Myb. Cell 131: 146-159. doi: 10.1016/j.cell.2007.07.021
|
| [9] |
He Y, Jiang X, Chen J (2014) The role of miR-150 in normal and malignant hematopoiesis. Oncogene 33: 3887-3893. doi: 10.1038/onc.2013.346
|
| [10] |
Guduric-Fuchs J, O'Connor A, Camp B, et al. (2012) Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genomics 13: 357. doi: 10.1186/1471-2164-13-357
|
| [11] |
de Candia P, Torri A, Gorletta T, et al. (2013) Intracellular modulation, extracellular disposal and serum increase of MiR-150 mark lymphocyte activation. PLoS One 8: e75348. doi: 10.1371/journal.pone.0075348
|
| [12] |
de Candia P, Torri A, Pagani M, et al. (2014) Serum microRNAs as biomarkers of human lymphocyte activation in health and disease. Front Immunol 5: 43. doi: 10.3389/fimmu.2014.00043
|
| [13] |
Oboshi W, Hayashi K, Takeuchi H, et al. (2020) MicroRNA-150 suppresses p27Kip1 expression and promotes cell proliferation in HeLa human cervical cancer cells. Oncol Lett 20: 210. doi: 10.3892/ol.2020.12073
|
| [14] |
Sur D, Burz C, Sabarimurugan S, et al. (2020) Diagnostic and prognostic significance of miR-150 in colorectal cancer: a systematic review and meta-analysis. J Pers Med 10: 99. doi: 10.3390/jpm10030099
|
| [15] |
Plank M, Maltby S, Mattes J, et al. (2013) Targeting translational control as a novel way to treat inflammatory disease: the emerging role of microRNAs. Clin Exp Allergy 43: 981-999. doi: 10.1111/cea.12135
|
| [16] |
Rebane A, Akdis CA (2014) MicroRNAs in allergy and asthma. Curr Allergy Asthma Rep 14: 424. doi: 10.1007/s11882-014-0424-x
|
| [17] |
Weidner J, Bartel S, Kılıç A, et al. (2020) Spotlight on microRNAs in allergy and asthma. Allergy 76: 1661-1678. doi: 10.1111/all.14646
|
| [18] |
Rebane A (2015) microRNA and allergy. Adv Exp Med Biol 888: 331-352. doi: 10.1007/978-3-319-22671-2_17
|
| [19] |
Garbacki N, Di Valentin E, Huynh-Thu VA, et al. (2011) MicroRNAs profiling in murine models of acute and chronic asthma: A relationship with mRNAs targets. PLoS One 6: e16509. doi: 10.1371/journal.pone.0016509
|
| [20] |
Feng MJ, Shi F, Qiu C, et al. (2012) MicroRNA-181a, -146a and -146b in spleen CD4+ T lymphocytes play proinflammatory roles in a murine model of asthma. Int Immunopharmacol 13: 347-353. doi: 10.1016/j.intimp.2012.05.001
|
| [21] | Badalzadeh M, Mazinani M, Pourpak Z, et al. (2019) In vitro analysis of nine microRNAs in CD8+ T cells of asthmatic patients and the effects of two FDA-approved drugs. Iran J Allergy Asthma Immunol 18: 358-368. |
| [22] |
Wang JW, Li K, Hellermann G, et al. (2012) MIR-150 suppresses lung inflammation in a mouse model of experimental asthma. World Allergy Organ J 5: S9. doi: 10.1097/01.WOX.0000411771.44473.bd
|
| [23] |
Wang JW, Li K, Hellermann G, et al. (2011) Regulating the regulators: microRNA and asthma. World Allergy Organ J 4: 94-103. doi: 10.1186/1939-4551-4-6-94
|
| [24] |
Zhang XY, Tang XY, Ma LJ, et al. (2017) Schisandrin B down-regulated lncRNA BCYRN1 expression of airway smooth muscle cells by improving miR-150 expression to inhibit the proliferation and migration of ASMC in asthmatic rats. Cell Proliferat 50: e12382. doi: 10.1111/cpr.12382
|
| [25] |
Zhang Q, Ni W, Li Y, et al. (2020) Analysis of altered miRNA profiling in the colon of a mouse model with β-lactoglobulin allergy. Allergol Immunopathol 48: 666-674. doi: 10.1016/j.aller.2020.05.007
|
| [26] |
Wąsik M, Nazimek K, Nowak B, et al. (2019) Delayed-type hypersensitivity underlying casein allergy is suppressed by extracellular vesicles carrying miR-150. Nutrients 11: 907. doi: 10.3390/nu11040907
|
| [27] |
Ho MHK, Wong WHS, Chang C (2014) Clinical spectrum of food allergies: a comprehensive review. Clin Rev Allergy Immu 46: 225-240. doi: 10.1007/s12016-012-8339-6
|
| [28] |
Vennegaard MT, Bonefeld CM, Hagedorn PH, et al. (2012) Allergic contact dermatitis induces upregulation of identical microRNAs in humans and mice. Contact Dermatitis 67: 298-305. doi: 10.1111/j.1600-0536.2012.02083.x
|
| [29] | Wolf J, Levis WR (2012) MicroRNA 150 in humans and murine contact sensitivity. J Drugs Dermatol 11: 1152. |
| [30] |
Zheng Q, Zhou L, Mi QS (2012) MicroRNA miR-150 is involved in Vα14 invariant NKT cell development and function. J Immunol 188: 2118-2126. doi: 10.4049/jimmunol.1103342
|
| [31] |
Goubier A, Vocanson M, Macari C, et al. (2013) Invariant NKT cells suppress CD8(+) T-cell-mediated allergic contact dermatitis independently of regulatory CD4(+) T cells. J Invest Dermatol 133: 980-987. doi: 10.1038/jid.2012.404
|
| [32] |
Askenase PW, Bryniarski K, Paliwal V, et al. (2015) A subset of AID-dependent B-1a cells initiates hypersensitivity and pneumococcal pneumonia resistance. Ann NY Acad Sci 1362: 200-214. doi: 10.1111/nyas.12975
|
| [33] |
Mi QS, Xu YP, Qi RQ, et al. (2012) Lack of microRNA miR-150 reduces the capacity of epidermal Langerhans cell cross-presentation. Exp Dermatol 21: 876-877. doi: 10.1111/exd.12008
|
| [34] |
Gulati N, Løvendorf MB, Zibert JR, et al. (2015) Unique microRNAs appear at different times during the course of a delayed-type hypersensitivity reaction in human skin. Exp Dermatol 24: 953-957. doi: 10.1111/exd.12813
|
| [35] |
Ptak W, Nazimek K, Askenase PW, et al. (2015) From mysterious supernatant entity to miR-150 in antigen-specific exosomes: a history of hapten-specific T suppressor factor. Arch Immunol Ther Exp 63: 345-356. doi: 10.1007/s00005-015-0331-4
|
| [36] |
Bryniarski K, Ptak W, Jayakumar A, et al. (2013) Antigen-specific, antibody-coated, exosome-like nanovesicles deliver suppressor T-cell microRNA-150 to effector T cells to inhibit contact sensitivity. J Allergy Clin Immun 132: 170-181. doi: 10.1016/j.jaci.2013.04.048
|
| [37] |
Nazimek K, Askenase PW, Bryniarski K (2018) Antibody light chains dictate the specificity of contact hypersensitivity effector cell suppression mediated by exosomes. Int J Mol Sci 19: 2656. doi: 10.3390/ijms19092656
|
| [38] |
Nazimek K, Bryniarski K, Ptak W, et al. (2020) Orally administered exosomes suppress mouse delayed-type hypersensitivity by delivering miR-150 to antigen-primed macrophage APC targeted by exosome-surface anti-peptide antibody light chains. Int J Mol Sci 21: 5540. doi: 10.3390/ijms21155540
|
| [39] |
Nazimek K, Ptak W, Nowak B, et al. (2015) Macrophages play an essential role in antigen-specific immune suppression mediated by T CD8+ cell-derived exosomes. Immunology 146: 23-32. doi: 10.1111/imm.12466
|
| [40] |
Nazimek K, Bustos-Morán E, Blas-Rus N, et al. (2019) Syngeneic red blood cell-induced extracellular vesicles suppress delayed-type hypersensitivity to self-antigens in mice. Clin Exp Allergy 49: 1487-1499. doi: 10.1111/cea.13475
|
| [41] | Nazimek K, Nowak B, Marcinkiewicz J, et al. (2014) Enhanced generation of reactive oxygen intermediates by suppressor T cell-derived exosome-treated macrophages. Folia Med Cracov 54: 37-52. |
| [42] |
Bryniarski K, Ptak W, Martin E, et al. (2015) Free extracellular miRNA functionally targets cells by transfecting exosomes from their companion cells. PLoS One 10: e0122991. doi: 10.1371/journal.pone.0122991
|
| [43] |
Huang XL, Zhang L, Li JP, et al. (2015) MicroRNA-150: A potential regulator in pathogens infection and autoimmune diseases. Autoimmunity 48: 503-510. doi: 10.3109/08916934.2015.1072518
|
| [44] |
Guan H, Peng R, Mao L, et al. (2020) Injured tubular epithelial cells activate fibroblasts to promote kidney fibrosis through miR-150-containing exosomes. Exp Cell Res 392: 112007. doi: 10.1016/j.yexcr.2020.112007
|
| [45] |
Luan J, Fu J, Chen C, et al. (2019) LNA-anti-miR-150 ameliorated kidney injury of lupus nephritis by inhibiting renal fibrosis and macrophage infiltration. Arthritis Res Ther 21: 276. doi: 10.1186/s13075-019-2044-2
|
| [46] |
Luan J, Fu J, Wang D, et al. (2020) miR-150-Based RNA interference attenuates tubulointerstitial fibrosis through the SOCS1/JAK/STAT pathway in vivo and in vitro. Mol Ther Nucleic Acids 22: 871-884. doi: 10.1016/j.omtn.2020.10.008
|
| [47] |
Zhou H, Hasni SA, Perez P, et al. (2013) miR-150 promotes renal fibrosis in lupus nephritis by downregulating SOCS1. J Am Soc Nephrol 24: 1073-1087. doi: 10.1681/ASN.2012080849
|
| [48] |
Du Z, Wu T, Liu L, et al. (2020) Extracellular vesicles-derived miR-150-5p secreted by adipose-derived mesenchymal stem cells inhibits CXCL1 expression to attenuate hepatic fibrosis. J Cell Mol Med 25: 701-715. doi: 10.1111/jcmm.16119
|
| [49] |
Honda N, Jinnin M, Kira-Etoh T, et al. (2013) miR-150 down-regulation contributes to the constitutive type I collagen overexpression in scleroderma dermal fibroblasts via the induction of integrin β3. Am J Pathol 182: 206-216. doi: 10.1016/j.ajpath.2012.09.023
|
| [50] |
Zidar N, Langner C, Jerala M, et al. (2020) Pathology of fibrosis in Crohn's disease-contribution to understanding its pathogenesis. Front Med 7: 167. doi: 10.3389/fmed.2020.00167
|
| [51] |
Ou H, Teng H, Qin Y, et al. (2020) Extracellular vesicles derived from microRNA-150-5p-overexpressing mesenchymal stem cells protect rat hearts against ischemia/reperfusion. Aging (Albany NY) 12: 12669-12683. doi: 10.18632/aging.102792
|
| [52] |
Yung S, Chan TM (2017) Molecular and immunological basis of tubulo-interstitial injury in lupus nephritis: a comprehensive review. Clin Rev Allergy Immu 52: 149-163. doi: 10.1007/s12016-016-8533-z
|
| [53] |
Okon LG, Werth VP (2013) Cutaneous lupus erythematosus: diagnosis and treatment. Best Pract Res Cl Rh 27: 391-404. doi: 10.1016/j.berh.2013.07.008
|
| [54] | Gensous N, Boizard-Moracchini A, Lazaro E, et al. (2020) Update on the cellular pathogenesis of lupus. Curr Opin Rheumatol In press. |
| [55] | Chang A, Clark MR, Ko K (2020) Cellular aspects of the pathogenesis of lupus nephritis. Curr Opin Rheumatol In press. |
| [56] |
Gorabi AM, Kiaie N, Aslani S, et al. (2020) Prospects for the potential of RNA interference in the treatment of autoimmune diseases: Small interfering RNAs in the spotlight. J Autoimmun 114: 102529. doi: 10.1016/j.jaut.2020.102529
|
| [57] |
Pauley KM, Cha S (2013) RNAi therapeutics in autoimmune disease. Pharmaceuticals (Basel) 6: 287-294. doi: 10.3390/ph6030287
|
| [58] |
Zhang H, Huang X, Ye L, et al. (2018) B cell-related circulating microRNAs with the potential value of biomarkers in the differential diagnosis, and distinguishment between the disease activity and lupus nephritis for systemic lupus erythematosus. Front Immunol 9: 1473. doi: 10.3389/fimmu.2018.01473
|
| [59] |
Steen SO, Iversen LV, Carlsen AL, et al. (2015) The circulating cell-free microRNA profile in systemic sclerosis is distinct from both healthy controls and systemic lupus erythematosus. J Rheumatol 42: 214-221. doi: 10.3899/jrheum.140502
|
| [60] | Nakhjavani M, Etemadi J, Pourlak T, et al. (2019) Plasma levels of miR-21, miR-150, miR-423 in patients with lupus nephritis. Iran J Kidney Dis 13: 198-206. |
| [61] | Su YJ, Tsai NW, Kung CT, et al. (2018) Investigation of microRNA in mitochondrial apoptotic pathway in systemic lupus erythematosus. Biomed Res Int 2018: 9026357. |
| [62] | Zhang M, Chen D, Zhang F, et al. (2020) Serum exosomal hsa-miR-135b-5p serves as a potential diagnostic biomarker in steroid-induced osteonecrosis of femoral head. Am J Transl Res 12: 2136-2154. |
| [63] |
Méndez-Flores S, Furuzawa-Carballeda J, Hernández-Molina G, et al. (2019) MicroRNA expression in cutaneous lupus: a new window to understand its pathogenesis. Mediators Inflamm 2019: 5049245. doi: 10.1155/2019/5049245
|
| [64] |
Wang H, Peng W, Ouyang X, et al. (2012) Circulating microRNAs as candidate biomarkers in patients with systemic lupus erythematosus. Transl Res 160: 198-206. doi: 10.1016/j.trsl.2012.04.002
|
| [65] |
Carlsen AL, Schetter AJ, Nielsen CT, et al. (2013) Circulating microRNA expression profiles associated with systemic lupus erythematosus. Arthritis Rheum 65: 1324-1334. doi: 10.1002/art.37890
|
| [66] |
Solé C, Moliné T, Vidal M, et al. (2019) An exosomal urinary miRNA signature for early diagnosis of renal fibrosis in lupus nephritis. Cells 8: 773. doi: 10.3390/cells8080773
|
| [67] |
Abulaban KM, Fall N, Nunna R, et al. (2016) Relationship of cell-free urine MicroRNA with lupus nephritis in children. Pediatric Rheumatol 14: 4. doi: 10.1186/s12969-016-0064-x
|
| [68] |
Omidi F, Hosseini SA, Ahmadi A, et al. (2020) Discovering the signature of a lupus-related microRNA profile in the Gene Expression Omnibus repository. Lupus 29: 1321-1335. doi: 10.1177/0961203320944473
|
| [69] |
Chen JQ, Papp G, Póliska S, et al. (2017) MicroRNA expression profiles identify disease-specific alterations in systemic lupus erythematosus and primary Sjögren's syndrome. PLoS One 12: e0174585. doi: 10.1371/journal.pone.0174585
|
| [70] |
Ebert MS, Sharp PA (2010) MicroRNA sponges: progress and possibilities. RNA 16: 2043-2050. doi: 10.1261/rna.2414110
|
| [71] |
Luan J, Jiao C, Kong W, et al. (2018) circHLA-C plays an important role in lupus nephritis by sponging miR-150. Mol Ther Nucleic Acids 10: 245-253. doi: 10.1016/j.omtn.2017.12.006
|
| [72] |
Lu MC, Lai NS, Chen HC, et al. (2013) Decreased microRNA(miR)-145 and increased miR-224 expression in T cells from patients with systemic lupus erythematosus involved in lupus immunopathogenesis. Clin Exp Immunol 171: 91-99. doi: 10.1111/j.1365-2249.2012.04676.x
|
| [73] |
Voynova EN, Skinner J, Bolland S (2015) Expansion of an atypical NK cell subset in mouse models of systemic lupus erythematosus. J Immunol 194: 1503-1513. doi: 10.4049/jimmunol.1402673
|
| [74] |
Gao S, Yuan L, Wang Y, et al. (2017) Enhanced expression of TREM-1 in splenic cDCs in lupus prone mice and it was modulated by miR-150. Mol Immunol 81: 127-134. doi: 10.1016/j.molimm.2016.12.006
|
| [75] |
Fox RI (2011) Extraglandular manifestations of Sjögren's Syndrome (SS): dermatologic arthritic endocrine, pulmonary, cardiovascular, gastroenterology, renal, urology, and gynecologic manifestations. Sjögren's Syndrome New York: Springer, 285-316. doi: 10.1007/978-1-60327-957-4_17
|
| [76] |
Lopes AP, Hillen MR, Chouri E, et al. (2018) Circulating small non-coding RNAs reflect IFN status and B cell hyperactivity in patients with primary Sjögren's syndrome. PLoS One 13: e0193157. doi: 10.1371/journal.pone.0193157
|
| [77] |
Ebrahimiyan H, Gharibdoost F, Aslani S, et al. (2020) microRNAs are potentially regulating the survivin gene in PBMCs from systemic sclerosis patients. Mod Rheumatol 30: 862-869. doi: 10.1080/14397595.2019.1659545
|
| [78] |
Heindryckx F, Binet F, Ponticos M, et al. (2016) Endoplasmic reticulum stress enhances fibrosis through IRE1α-mediated degradation of miR-150 and XBP-1 splicing. EMBO Mol Med 8: 729-744. doi: 10.15252/emmm.201505925
|
| [79] |
Jinnin M (2014) Various applications of microRNAs in skin diseases. J Dermatol Sci 74: 3-8. doi: 10.1016/j.jdermsci.2014.01.004
|
| [80] |
Luo Y, Xiao R (2020) The epigenetic regulation of scleroderma and its clinical application. Adv Exp Med Biol 1253: 375-403. doi: 10.1007/978-981-15-3449-2_13
|
| [81] |
Page A, Fusil F, Cosset FL (2021) Antigen-specific tolerance approach for rheumatoid arthritis: past, present and future. Joint Bone Spine 88: 105164. doi: 10.1016/j.jbspin.2021.105164
|
| [82] |
Churov AV, Oleinik EK, Knip M (2015) MicroRNAs in rheumatoid arthritis: altered expression and diagnostic potential. Autoimmun Rev 14: 1029-1037. doi: 10.1016/j.autrev.2015.07.005
|
| [83] |
Qiu M, Mo L, Li J, et al. (2020) Effects of miR-150-5p on the growth and SOCS1 expression of rheumatoid arthritis synovial fibroblasts. Clin Rheumatol 39: 909-917. doi: 10.1007/s10067-019-04894-7
|
| [84] |
Zhao F, Dong J, Guo J, et al. (2020) Inhibiting role of long non-coding RNA LINC01197 in inflammation in rheumatoid arthritis through the microRNA-150/THBS2 axis. Exp Cell Res 394: 112136. doi: 10.1016/j.yexcr.2020.112136
|
| [85] |
Niimoto T, Nakasa T, Ishikawa M, et al. (2010) MicroRNA-146a expresses in interleukin-17 producing T cells in rheumatoid arthritis patients. BMC Musculoskelet Disord 11: 209. doi: 10.1186/1471-2474-11-209
|
| [86] |
Ebrahimiyan H, Rezaei N, Vojdanian M, et al. (2019) microRNA involvement in the regulation of survivin in peripheral blood mononuclear cells from rheumatoid arthritis patients. Int J Rheum Dis 22: 1107-1114. doi: 10.1111/1756-185X.13520
|
| [87] |
Rezaeepoor M, Pourjafar M, Tahamoli-Roudsari A, et al. (2020) Altered expression of microRNAs may predict therapeutic response in rheumatoid arthritis patients. Int Immunopharmacol 83: 106404. doi: 10.1016/j.intimp.2020.106404
|
| [88] |
Anaparti V, Smolik I, Meng X, et al. (2017) Whole blood microRNA expression pattern differentiates patients with rheumatoid arthritis, their seropositive first-degree relatives, and healthy unrelated control subjects. Arthritis Res Ther 19: 249. doi: 10.1186/s13075-017-1459-x
|
| [89] |
Chen Z, Wang H, Xia Y, et al. (2018) Therapeutic potential of mesenchymal cell-derived miR-150-5p-expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J Immunol 201: 2472-2482. doi: 10.4049/jimmunol.1800304
|
| [90] |
Elzorkany B, Mokbel A, Gamal SM, et al. (2021) Does smoking affect level of seropositivity in RA? A post-HOC global and inter-country analysis of COMORA cohort. Rheumatol Int 41: 699-705. doi: 10.1007/s00296-021-04791-w
|
| [91] |
Wasén C, Ospelt C, Camponeschi A, et al. (2020) Nicotine changes the microRNA profile to regulate the FOXO memory program of CD8+ T cells in rheumatoid arthritis. Front Immunol 11: 1474. doi: 10.3389/fimmu.2020.01474
|
| [92] |
Magrey MN, Haqqi T, Haseeb A (2016) Identification of plasma microRNA expression profile in radiographic axial spondyloarthritis-a pilot study. Clin Rheumatol 35: 1323-1327. doi: 10.1007/s10067-015-3123-7
|
| [93] |
Perez-Sanchez C, Font-Ugalde P, Ruiz-Limon P, et al. (2018) Circulating microRNAs as potential biomarkers of disease activity and structural damage in ankylosing spondylitis patients. Hum Mol Genet 27: 875-890. doi: 10.1093/hmg/ddy008
|
| [94] |
Lerman G, Avivi C, Mardoukh C, et al. (2011) MiRNA expression in psoriatic skin: reciprocal regulation of hsa-miR-99a and IGF-1R. PLoS One 6: e20916. doi: 10.1371/journal.pone.0020916
|
| [95] |
Li Y, Su J, Li F, et al. (2017) MiR-150 regulates human keratinocyte proliferation in hypoxic conditions through targeting HIF-1α and VEGFA: Implications for psoriasis treatment. PLoS One 12: e0175459. doi: 10.1371/journal.pone.0175459
|
| [96] |
Zhu WJ, Li P, Wang L, et al. (2020) Hypoxia-inducible factor-1: A potential pharmacological target to manage psoriasis. Int Immunopharmacol 86: 106689. doi: 10.1016/j.intimp.2020.106689
|
| [97] |
Torri A, Carpi D, Bulgheroni E, et al. (2017) Extracellular microRNA signature of human helper T cell subsets in health and autoimmunity. J Biol Chem 292: 2903-2915. doi: 10.1074/jbc.M116.769893
|
| [98] |
Okoye IS, Coomes SM, Pelly VS, et al. (2014) MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity 41: 89-103. doi: 10.1016/j.immuni.2014.05.019
|
| [99] |
Chen WX, Ren LH, Shi RH (2014) Implication of miRNAs for inflammatory bowel disease treatment: Systematic review. World J Gastrointest Pathophysiol 5: 63-70. doi: 10.4291/wjgp.v5.i2.63
|
| [100] |
Bian Z, Li L, Cui J, et al. (2011) Role of miR-150-targeting c-Myb in colonic epithelial disruption during dextran sulphate sodium-induced murine experimental colitis and human ulcerative colitis. J Pathol 225: 544-553. doi: 10.1002/path.2907
|
| [101] |
Rodríguez-Nogales A, Algieri F, Garrido-Mesa J, et al. (2018) The administration of Escherichia coli Nissle 1917 ameliorates development of DSS-induced colitis in mice. Front Pharmacol 9: 468. doi: 10.3389/fphar.2018.00468
|
| [102] |
Din AU, Hassan A, Zhu Y, et al. (2020) Inhibitory effect of Bifidobacterium bifidum ATCC 29521 on colitis and its mechanism. J Nutr Biochem 79: 108353. doi: 10.1016/j.jnutbio.2020.108353
|
| [103] |
Rodríguez-Nogales A, Algieri F, Garrido-Mesa J, et al. (2017) Differential intestinal anti-inflammatory effects of Lactobacillus fermentum and Lactobacillus salivarius in DSS mouse colitis: impact on microRNAs expression and microbiota composition. Mol Nutr Food Res 61: 1700144. doi: 10.1002/mnfr.201700144
|
| [104] |
Morris NL, Hammer AM, Cannon AR, et al. (2017) Dysregulation of microRNA biogenesis in the small intestine after ethanol and burn injury. BBA-Mol Basis Dis 1863: 2645-2653. doi: 10.1016/j.bbadis.2017.03.025
|
| [105] | Wang S, Huang Y, Zhou C, et al. (2018) The role of autophagy and related microRNAs in inflammatory bowel disease. Gastroent Res Pract 2018: 7565076. |
| [106] |
Ciccacci C, Politi C, Novelli G, et al. (2016) Advances in exploring the role of microRNAs in inflammatory bowel disease. MicroRNA 5: 5-11. doi: 10.2174/2211536605666160111124812
|
| [107] | Luo J, Wang Y, Lan D, et al. (2018) Differential expression of serum microRNAs in glucocorticoid-resistant patients with ulcerative colitis. Int J Clin Exp Pathol 11: 936-946. |
| [108] |
Bao Y, Guo Y, Li Z, et al. (2014) MicroRNA profiling in Muc2 knockout mice of colitis-associated cancer model reveals epigenetic alterations during chronic colitis malignant transformation. PLoS One 9: e99132. doi: 10.1371/journal.pone.0099132
|
| [109] |
Zamvil SS, Hauser SL (2021) Antigen presentation by B cells in multiple sclerosis. N Engl J Med 384: 378-381. doi: 10.1056/NEJMcibr2032177
|
| [110] |
Hu Z, Cui Y, Qiao X, et al. (2018) Silencing miR-150 ameliorates experimental autoimmune encephalomyelitis. Front Neurosci 12: 465. doi: 10.3389/fnins.2018.00465
|
| [111] |
Fenoglio C, Cantoni C, Riz MD, et al. (2011) Expression and genetic analysis of miRNAs involved in CD4+ cell activation in patients with multiple sclerosis. Neurosci Lett 504: 9-12. doi: 10.1016/j.neulet.2011.08.021
|
| [112] |
Martinelli-Boneschi F, Fenoglio C, Brambilla P, et al. (2012) MicroRNA and mRNA expression profile screening in multiple sclerosis patients to unravel novel pathogenic steps and identify potential biomarkers. Neurosci Lett 508: 4-8. doi: 10.1016/j.neulet.2011.11.006
|
| [113] |
Jernås M, Malmeström C, Axelsson M, et al. (2013) MicroRNA regulate immune pathways in T-cells in multiple sclerosis (MS). BMC Immunol 14: 32. doi: 10.1186/1471-2172-14-32
|
| [114] |
Bergman P, Piket E, Khademi M, et al. (2016) Circulating miR-150 in CSF is a novel candidate biomarker for multiple sclerosis. Neurol Neuroimmunol Neuroinflamm 3: e219. doi: 10.1212/NXI.0000000000000219
|
| [115] |
Quintana E, Ortega FJ, Robles-Cedeño R, et al. (2017) miRNAs in cerebrospinal fluid identify patients with MS and specifically those with lipid-specific oligoclonal IgM bands. Mult Scler 23: 1716-1726. doi: 10.1177/1352458516684213
|
| [116] |
Bruinsma IB, van Dijk M, Bridel C, et al. (2017) Regulator of oligodendrocyte maturation, miR-219, a potential biomarker for MS. J Neuroinflammation 14: 235. doi: 10.1186/s12974-017-1006-3
|
| [117] |
Shakerian L, Ghorbani S, Talebi F, et al. (2018) MicroRNA-150 targets PU.1 and regulates macrophage differentiation and function in experimental autoimmune encephalomyelitis. J Neuroimmunol 323: 167-174. doi: 10.1016/j.jneuroim.2018.06.010
|
| [118] |
Dolati S, Aghebati-Maleki L, Ahmadi M, et al. (2018) Nanocurcumin restores aberrant miRNA expression profile in multiple sclerosis, randomized, double-blind, placebo-controlled trial. J Cell Physiol 233: 5222-5230. doi: 10.1002/jcp.26301
|
| [119] |
Al-Ghezi ZZ, Miranda K, Nagarkatti M, et al. (2019) Combination of cannabinoids, Δ9-tetrahydrocannabinol and cannabidiol, ameliorates experimental multiple sclerosis by suppressing neuroinflammation through regulation of miRNA-mediated signaling pathways. Front Immunol 10: 1921. doi: 10.3389/fimmu.2019.01921
|
| [120] |
Nadin T, Haque A, Akil M, et al. (2019) Management of the idiopathic inflammatory myopathies. Prescriber 30: 28-33. doi: 10.1002/psb.1762
|
| [121] |
Cotton T, Niaki OZ, Zheng B, et al. (2021) Myositis in systemic lupus erythematosus. Lupus 30: 615-619. doi: 10.1177/0961203320988587
|
| [122] | Ye L, Zuo Y, Yang H, et al. (2019) Specific autoantibodies and clinical phenotypes correlate with the aberrant expression of immune-related microRNAs in dermatomyositis. J Immunol Res 2019: 2927061. |
| [123] |
Punga T, Le Panse R, Andersson M, et al. (2014) Circulating miRNAs in myasthenia gravis: miR-150-5p as a new potential biomarker. Ann Clin Transl Neurol 1: 49-58. doi: 10.1002/acn3.24
|
| [124] |
Punga AR, Andersson M, Alimohammadi M, et al. (2015) Disease specific signature of circulating miR-150-5p and miR-21-5p in myasthenia gravis patients. J Neurol Sci 356: 90-96. doi: 10.1016/j.jns.2015.06.019
|
| [125] |
Molin CJ, Sabre L, Weis CA, et al. (2018) Thymectomy lowers the myasthenia gravis biomarker miR-150-5p. Neurol Neuroimmunol Neuroinflamm 5: e450. doi: 10.1212/NXI.0000000000000450
|
| [126] |
Westerberg E, Molin CJ, Lindblad I, et al. (2017) Physical exercise in myasthenia gravis is safe and improves neuromuscular parameters and physical performance-based measures: A pilot study. Muscle Nerve 56: 207-214. doi: 10.1002/mus.25493
|
| [127] |
Sabre L, Maddison P, Sadalage G, et al. (2018) Circulating microRNA miR-21-5p, miR-150-5p and miR-30e-5p correlate with clinical status in late onset myasthenia gravis. J Neuroimmunol 321: 164-170. doi: 10.1016/j.jneuroim.2018.05.003
|
| [128] |
Punga AR, Punga T (2018) Circulating microRNAs as potential biomarkers in myasthenia gravis patients. Ann NY Acad Sci 1412: 33-40. doi: 10.1111/nyas.13510
|
| [129] |
Sabre L, Punga T, Punga AR (2020) Circulating miRNAs as potential biomarkers in myasthenia gravis: tools for personalized medicine. Front Immunol 11: 213. doi: 10.3389/fimmu.2020.00213
|
| [130] |
Sabre L, Maddison P, Wong SH, et al. (2019) miR-30e-5p as predictor of generalization in ocular myasthenia gravis. Ann Clin Transl Neurol 6: 243-251. doi: 10.1002/acn3.692
|
| [131] |
Fiorillo AA, Heier CR, Huang YF, et al. (2020) Estrogen receptor, inflammatory, and FOXO transcription factors regulate expression of myasthenia gravis-associated circulating microRNAs. Front Immunol 11: 151. doi: 10.3389/fimmu.2020.00151
|
| [132] |
Zhong H, Lu J, Jing S, et al. (2020) Low-dose rituximab lowers serum Exosomal miR-150-5p in AChR-positive refractory myasthenia gravis patients. J Neuroimmunol 348: 577383. doi: 10.1016/j.jneuroim.2020.577383
|
| [133] |
Cron MA, Maillard S, Truffault F, et al. (2019) Causes and consequences of miR-150-5p dysregulation in myasthenia gravis. Front Immunol 10: 539. doi: 10.3389/fimmu.2019.00539
|
| [134] |
Cron MA, Guillochon É, Kusner L, et al. (2020) Role of miRNAs in normal and myasthenia gravis thymus. Front Immunol 11: 1074. doi: 10.3389/fimmu.2020.01074
|
| [135] |
Ke Q, Kroger CJ, Clark M, et al. (2020) Evolving antibody therapies for the treatment of type 1 diabetes. Front Immunol 11: 624568. doi: 10.3389/fimmu.2020.624568
|
| [136] |
Estrella S, Garcia-Diaz DF, Codner E, et al. (2016) Expression of miR-22 and miR-150 in type 1 diabetes mellitus: Possible relationship with autoimmunity and clinical characteristics. Med Clin-Barcelona 147: 245-247. doi: 10.1016/j.medcli.2016.05.016
|
| [137] |
Wang G, Gu Y, Xu N, et al. (2018) Decreased expression of miR-150, miR146a and miR424 in type 1 diabetic patients: Association with ongoing islet autoimmunity. Biochem Biophys Res Commun 498: 382-387. doi: 10.1016/j.bbrc.2017.06.196
|
| [138] |
Assmann TS, Recamonde-Mendoza M, De Souza BM, et al. (2017) MicroRNA expression profiles and type 1 diabetes mellitus: systematic review and bioinformatic analysis. Endocr Connect 6: 773-790. doi: 10.1530/EC-17-0248
|
| [139] |
Mazzeo A, Beltramo E, Lopatina T, et al. (2018) Molecular and functional characterization of circulating extracellular vesicles from diabetic patients with and without retinopathy and healthy subjects. Exp Eye Res 176: 69-77. doi: 10.1016/j.exer.2018.07.003
|
| [140] |
Kim H, Bae YU, Jeon JS, et al. (2019) The circulating exosomal microRNAs related to albuminuria in patients with diabetic nephropathy. J Transl Med 17: 236. doi: 10.1186/s12967-019-1983-3
|
| [141] |
Lee WC, Li LC, Ng HY, et al. (2020) Urinary exosomal microRNA signatures in nephrotic, biopsy-proven diabetic nephropathy. J Clin Med 9: 1220. doi: 10.3390/jcm9041220
|
| [142] |
Mazzeo A, Lopatina T, Gai C, et al. (2019) Functional analysis of miR-21-3p, miR-30b-5p and miR-150-5p shuttled by extracellular vesicles from diabetic subjects reveals their association with diabetic retinopathy. Exp Eye Res 184: 56-63. doi: 10.1016/j.exer.2019.04.015
|
| [143] |
Henriques-Antunes H, Cardoso RMS, Zonari A, et al. (2019) The kinetics of small extracellular vesicle delivery impacts skin tissue regeneration. ACS Nano 13: 8694-8707. doi: 10.1021/acsnano.9b00376
|
| [144] |
Tian J, Pan W, Xu X, et al. (2020) NF-κB inhibits the occurrence of type 1 diabetes through microRNA-150-dependent PUMA degradation. Life Sci 255: 117724. doi: 10.1016/j.lfs.2020.117724
|
| [145] |
Roat R, Hossain MM, Christopherson J, et al. (2019) Circulating miR-150-5p is associated with immune-mediated early β-cell loss in a humanized mouse model. Xenotransplantation 26: e12474. doi: 10.1111/xen.12474
|
| [146] |
Hamada S, Masamune A, Kanno A, et al. (2015) Comprehensive analysis of serum microRNAs in autoimmune pancreatitis. Digestion 91: 263-271. doi: 10.1159/000381283
|
| [147] |
Wasik U, Kempinska-Podhorodecka A, Bogdanos DP, et al. (2020) Enhanced expression of miR-21 and miR-150 is a feature of anti-mitochondrial antibody-negative primary biliary cholangitis. Mol Med 26: 8. doi: 10.1186/s10020-019-0130-1
|
| [148] |
Xing L, Xu W, Qu Y, et al. (2018) miR-150 regulates B lymphocyte in autoimmune hemolytic anemia/Evans syndrome by c-Myb. Int J Hematol 107: 666-672. doi: 10.1007/s12185-018-2429-z
|
| [149] |
Lucherini OM, Obici L, Ferracin M, et al. (2013) First report of circulating microRNAs in tumour necrosis factor receptor-associated periodic syndrome (TRAPS). PLoS One 8: e73443. doi: 10.1371/journal.pone.0073443
|
| [150] | Nazimek K, Filipczak-Bryniarska I, Bryniarski K (2015) The role of medicaments, exosomes and miRNA molecules in modulation of macrophage immune activity. Postepy Hig Med Dosw 69: 1114-1129. |
| [151] |
Nazimek K, Bryniarski K (2012) The biological activity of macrophages in health and disease. Postepy Hig Med Dosw 66: 507-520. doi: 10.5604/17322693.1004080
|
| [152] |
Trifari S, Pipkin ME, Bandukwala HS, et al. (2013) MicroRNA-directed program of cytotoxic CD8+ T-cell differentiation. P Natl Acad Sci USA 110: 18608-18613. doi: 10.1073/pnas.1317191110
|
| [153] |
Liang Y, Pan HF, Ye DQ (2015) microRNAs function in CD8+ T cell biology. J Leukocyte Biol 97: 487-497. doi: 10.1189/jlb.1RU0814-369R
|
| [154] |
Ma Z, Shen Y, Zeng Q, et al. (2018) MiR-150-5p regulates EGR2 to promote the development of chronic rhinosinusitis via the DC-Th axis. Int Immunopharmacol 54: 188-197. doi: 10.1016/j.intimp.2017.11.011
|
| [155] | Nazimek K, Bryniarski K, Santocki M, et al. (2015) Exosomes as mediators of intercellular communication: clinical implications. Pol Arch Med Wewn 125: 370-380. |
| [156] |
Sang W, Sun C, Zhang C, et al. (2016) MicroRNA-150 negatively regulates the function of CD4(+) T cells through AKT3/Bim signaling pathway. Cell Immunol 306–307: 35-40. doi: 10.1016/j.cellimm.2016.05.007
|
| [157] |
Sang W, Wang Y, Zhang C, et al. (2016) MiR-150 impairs inflammatory cytokine production by targeting ARRB-2 after blocking CD28/B7 costimulatory pathway. Immunol Lett 172: 1-10. doi: 10.1016/j.imlet.2015.11.001
|
| [158] |
Tung SL, Boardman DA, Sen M, et al. (2018) Regulatory T cell-derived extracellular vesicles modify dendritic cell function. Sci Rep 8: 6065. doi: 10.1038/s41598-018-24531-8
|
| [159] |
Neamah WH, Singh NP, Alghetaa H, et al. (2019) AhR activation leads to massive mobilization of myeloid-derived suppressor cells with immunosuppressive activity through regulation of CXCR2 and microRNA miR-150-5p and miR-543-3p that target anti-inflammatory genes. J Immunol 203: 1830-1844. doi: 10.4049/jimmunol.1900291
|
| [160] |
Warth SC, Hoefig KP, Hiekel A, et al. (2015) Induced miR-99a expression represses Mtor cooperatively with miR-150 to promote regulatory T-cell differentiation. EMBO J 34: 1195-1213. doi: 10.15252/embj.201489589
|
| [161] |
Kluiver JL, Chen C-Z (2012) MicroRNAs regulate B-cell receptor signaling-induced apoptosis. Genes Immun 13: 239-244. doi: 10.1038/gene.2012.1
|
| [162] |
Mraz M, Chen L, Rassenti LZ, et al. (2014) miR-150 influences B-cell receptor signaling in chronic lymphocytic leukemia by regulating expression of GAB1 and FOXP1. Blood 124: 84-95. doi: 10.1182/blood-2013-09-527234
|
| [163] |
Cerna K, Oppelt J, Chochola V, et al. (2019) MicroRNA miR-34a downregulates FOXP1 during DNA damage response to limit BCR signalling in chronic lymphocytic leukaemia B cells. Leukemia 33: 403-414. doi: 10.1038/s41375-018-0230-x
|
| [164] |
Musilova K, Devan J, Cerna K, et al. (2018) miR-150 downregulation contributes to the high-grade transformation of follicular lymphoma by upregulating FOXP1 levels. Blood 132: 2389-2400. doi: 10.1182/blood-2018-06-855502
|
| [165] |
Jiang XX, Liu Y, Li H, et al. (2016) MYSM1/miR-150/FLT3 inhibits B1a cell proliferation. Oncotarget 7: 68086-68096. doi: 10.18632/oncotarget.11738
|
| [166] |
Ma Y, Liu Y, Hou H, et al. (2018) MiR-150 predicts survival in patients with sepsis and inhibits LPS-induced inflammatory factors and apoptosis by targeting NF-κB1 in human umbilical vein endothelial cells. Biochem Biophys Res Commun 500: 828-837. doi: 10.1016/j.bbrc.2018.04.168
|
| [167] |
Palagani A, Op de Beeck K, Naulaerts S, et al. (2014) Ectopic microRNA-150-5p transcription sensitizes glucocorticoid therapy response in MM1S multiple myeloma cells but fails to overcome hormone therapy resistance in MM1R cells. PLoS One 9: e113842. doi: 10.1371/journal.pone.0113842
|
| [168] |
Nazimek K, Bustos-Morán E, Blas-Rus N, et al. (2021) Antibodies enhance the suppressive activity of extracellular vesicles in mouse delayed-type hypersensitivity. Pharmaceuticals 14: 734. doi: 10.3390/ph14080734
|


Figures(3) / Tables(1)

Katarzyna Nazimek. The complex functions of microRNA-150 in allergy, autoimmunity and immune tolerance[J]. AIMS Allergy and Immunology, 2021, 5(4): 195-221. doi: 10.3934/Allergy.2021016







 DownLoad:
DownLoad: