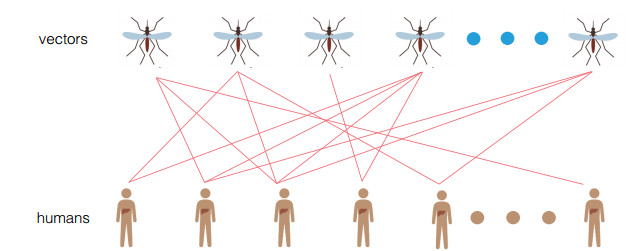
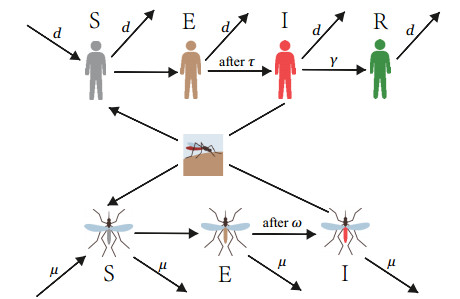
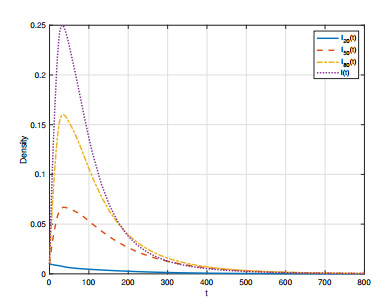
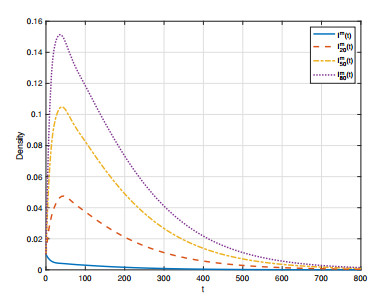
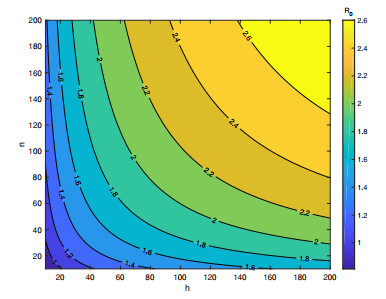
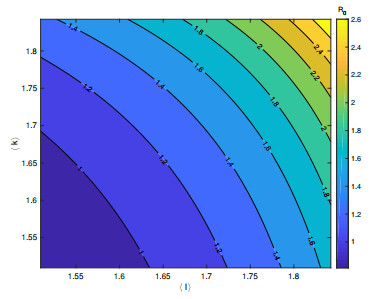
In order to study the impact of the incubation periods of humans and vectors on diseases transmission, a novel vector-borne diseases model with two time delays on bipartite networks is proposed. The formula of the basic reproduction number $ R_0 $ is given, which is dependent on time delays. Moreover, the globally asymptotic stability of the disease-free equilibrium and the endemic equilibrium is proved by constructing appropriate Lyapunov functions. Finally, numerical simulations are carried out to verify the analysis results and reveal the influence of the structure of bipartite networks on the basic reproduction number.
Citation: Rundong Zhao, Qiming Liu, Huazong Zhang. Dynamical behaviors of a vector-borne diseases model with two time delays on bipartite networks[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3073-3091. doi: 10.3934/mbe.2021154
In order to study the impact of the incubation periods of humans and vectors on diseases transmission, a novel vector-borne diseases model with two time delays on bipartite networks is proposed. The formula of the basic reproduction number $ R_0 $ is given, which is dependent on time delays. Moreover, the globally asymptotic stability of the disease-free equilibrium and the endemic equilibrium is proved by constructing appropriate Lyapunov functions. Finally, numerical simulations are carried out to verify the analysis results and reveal the influence of the structure of bipartite networks on the basic reproduction number.

| [1] | S. M. Lemon, P. F. Sparling, M. A. Hamburg, D. A. Relman, E. R. Choffnes, Vector-borne diseases: Understanding the environmental, human health, and ecological connections. Workshop summary, National Academies Press, 2008. |
| [2] | Changes in the incidence of vector-borne disease attributable to climate change, CIESIN, 2007. Available from: http://www.ciesin.columbia.edu/TG/HH/veclev2. |
| [3] | R. Ross, The prevention of malaria, John Murray, 1911. |
| [4] |
A. Traoré, Analysis of a vector-borne disease model with human and vectors immigration, J. Appl. Math. Comput., 64 (2020), 411–428. doi: 10.1007/s12190-020-01361-4

|
| [5] | R. Nyang'inja, G. Lawi, M. Okongo, A. Orwa, Stability analysis of Rotavirus-malaria co-epidemic model with vaccination, Dyn. Syst. Appl., 28 (2019), 371–407. |
| [6] | A. Abiodun, P. Witbooi, K. O. Okosun, Modelling the impact of climatic variables on malaria transmission, Hacet. J. Math. Stat., 47 (2018), 219–235. |
| [7] |
A. L. Barabási, R. Albert, Emergence of scaling in random networks, Science, 286 (1999), 509–512. doi: 10.1126/science.286.5439.509
|
| [8] |
S. Boccaletti, V. Latora, Y. Moreno, M. Chavez, D. U. Hwang, Complex networks: Structure and dynamics, Phys. Rep., 424 (2006), 175–308. doi: 10.1016/j.physrep.2005.10.009
|
| [9] |
S. Eubank, H. Guclu, V. S. Anil Kumar, M. V. Marathe, A. Srinivasan, Z. Toroczkai et al., Modelling disease outbreaks in realistic urban social networks, Nature, 429 (2004), 180–184. doi: 10.1038/nature02541
|
| [10] |
N. Masuda, N. Konno, Multi-state epidemic processes on complex networks, J. Theor. Biol., 243 (2006), 64–75. doi: 10.1016/j.jtbi.2006.06.010
|
| [11] |
H. Shi, Z. Duan, G. Chen, An SIS model with infective medium on complex networks, Phys. A, 387 (2008), 2133–2144. doi: 10.1016/j.physa.2007.11.048
|
| [12] |
M. E. Woolhouse, C. Dye, J.-F. Etard, T. Smith, J. Charlwood, G. Garnett, et al., Heterogeneities in the transmission of infectious agents: Implications for the design of control programs, Proc. Natl. Acad. Sci. U.S.A., 94 (1997), 338–342. doi: 10.1073/pnas.94.1.338
|
| [13] |
M. Yang, G. Chen, X. Fu, A modfied SIS model with an infective medium on complex networks and its global stability, Phys. A, 390 (2011), 2408–2413. doi: 10.1016/j.physa.2011.02.007
|
| [14] |
Y. Wang, Z. Jin, Z. Yang, Z. Zhang, T. Zhou, G. Sun, Global analysis of an SIS model with an infective vector on complex networks, Nonlinear Anal.-Real World Appl., 13 (2012), 543–557. doi: 10.1016/j.nonrwa.2011.07.033
|
| [15] |
X. Wang, J. Yang, Dynamical analysis of a mean-field vector-borne diseases model on complex networks: An edge based compartmental approach, Chaos, 30 (2020), 013103. doi: 10.1063/1.5116209
|
| [16] |
R. Zhang, Global dynamic analysis of a model for vector-borne diseases on bipartite networks, Phys. A, 545 (2020), 123813. doi: 10.1016/j.physa.2019.123813
|
| [17] |
R. Bray, P. Garnham, The life-cycle of primate malaria parasites, Br. Med. Bull., 38 (1982), 117–122. doi: 10.1093/oxfordjournals.bmb.a071746
|
| [18] | D. M. Claborn, P. M. Masuoka, T. A. Klein, T. Hooper, A. Lee, R. G. Andre, A cost comparison of two malaria control methods in Kyunggi province, Republic of Korea, using remote sensing and geographic information systems. Am. J. Trop. Med. Hyg., 66 (2002), 680–685. |
| [19] | R. Zhang, D. Li, Z. Jin, Dynamic analysis of a delayed model for vector-borne diseases on bipartite networks, Appl. Math. Comput., 263 (2015), 342–352. |
| [20] |
C. Li, C. Tsai, S. Yang, Analysis of epidemic spreading of an SIRS model in complex heterogeneous networks, Commun. Nonlinear Sci. Numer. Simul., 19 (2014), 1042–1054. doi: 10.1016/j.cnsns.2013.08.033
|
| [21] |
J. Liu, T. Zhang, Epidemic spreading of an SEIRS model in scale-free networks, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 3375–3384. doi: 10.1016/j.cnsns.2010.11.019
|
| [22] |
H. Zhang, X. Fu, Spreading of epidemics on scale-free networks with nonlinear infectivity, Nonlinear Anal.-Theory Methods Appl., 70 (2009), 3273–3278. doi: 10.1016/j.na.2008.04.031
|
| [23] | J. K. Hale, S. M. V. Lunel, L. S. Verduyn, S. M. V. Lunel, Introduction to Functional Differential Equations, vol. 99, Springer Science & Business Media1 1993. |
| [24] | J. P. La Salle, The Stability of Dynamical Systems, SIAM, 1976. |
| [25] |
L. L. Shapiro, S. A. Whitehead, M. B. Thomas, Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria, PLoS. Biol., 15 (2017), e2003489. doi: 10.1371/journal.pbio.2003489
|
| [26] |
S. Ruan, D. Xiao, J. C. Beier, On the delayed Ross-Macdonald model for malaria transmission, Bull. Math. Biol., 70 (2008), 1098–1114. doi: 10.1007/s11538-007-9292-z
|
| [27] | Global health estimates: Life expectancy and leading causes of death and disability, W. H. Organization, 2021. Available from: https://www.who.int/data/gho/data/indicators/indicator-details/GHO/gho-ghe-halehealthy-life-expectancy-at-birth. |
| [28] | Malaria, W. H. Organization, 2021. Available from: https://www.who.int/health-topics/malaria#tab=tab_1. |
Figures(14) / Tables(2)

Rundong Zhao, Qiming Liu, Huazong Zhang. Dynamical behaviors of a vector-borne diseases model with two time delays on bipartite networks[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3073-3091. doi: 10.3934/mbe.2021154







 DownLoad:
DownLoad: