Citation: Yanmei Wang, Guirong Liu. Dynamics analysis of a stochastic SIRS epidemic model with nonlinear incidence rate and transfer from infectious to susceptible[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 6047-6070. doi: 10.3934/mbe.2019303

| [1] | F. H. Chen, A susceptible-infected epidemic model with voluntary vaccinations, J. Math. Biol., 53(2006), 253–272. |
| [2] | J. Li and Z. Ma, Stability analysis for SIS epidemic models with vaccination and constant popu-lation size, Discrete Contin. Dyn. Syst., Ser. B, 4(2004), 635–642. |
| [3] | F. Zhang, J. Li and J. Li, Epidemic characteristics of two classic SIS models with disease-induced death, J. Theor. Biol., 424(2017), 73–83. |
| [4] | W. M. Liu, H. W. Hethcote and S. A. Levin, Dynamical behavior of epidemiological models with nonlinear incidence rates, J. Math. Biol., 25(1987), 359–380. |
| [5] | W. M. Liu, S. A. Levin and Y. Iwasa, Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models, J. Math. Biol., 23(1986), 187–204. |
| [6] | T. Li, F. Zhang, H. Liu, et al., Threshold dynamics of an SIRS model with nonlinear incidence rate and transfer from infectious to susceptible, Appl. Math. Lett., 70(2017), 52–57. |
| [7] | T. Khan, A. Khan and G. Zaman, The extinction and persistence of the stochastic hepatitis B epidemic model, Chaos Soliton Fract., 108(2018), 123–128. |
| [8] | X. Meng, F. Li and S. Gao, Global analysis and numerical simulations of a novel stochastic eco-epidemiological model with time delay, Appl. Math. Comput., 339(2018), 701–726. |
| [9] | M. EI Fatini, A. Lahrouz, R. Pettersson, et al., Stochstic stability and instability of an epidemic model with relapse, Appl. Math. Comput., 316(2018), 326–341. |
| [10] | A. Lahrouz and A. Settati, Necessary and sufficient condition for extinction and persistence of SIRS system with random perturbation, Appl, Math, Comput., 233(2014), 10–19. |
| [11] | Y. Lin and D. Jiang, Threshold behavior in a stochastic SIS epidemic model with standard inci-dence, J. Dyn. Diff. Equat., 26(2014), 1079–1094. |
| [12] | Z. Teng and L. Wang, Persistence and extinction for a class of stochastic SIS epidemic models with nonlinear incidence rate, Physic. A, 451(2016), 507–518. |
| [13] | H. Qi, L. Liu and X. Meng, Dynamics of a non-autonomous stochastic SIS epidemic model with double epidemic hypothesis, Complexity, 2017(2017), 14. |
| [14] | Y. Cai, Y. Kang and W. Wang, A stochastic SIRS epidemic model with nonlinear incidence rate, Appl. Math. Comput., 305(2017), 221–240. |
| [15] | Z. Chang, X. Meng and X. Lu, Analysis of a novel stochastic SIRS epidemic model with two different saturated incidence rates, Physic. A, 472(2017), 103–116. |
| [16] | N. H. Du and N. N. Nhu, Permanence and extinction of certain stochastic SIR models perturbed by a complex type of noises, Appl. Math. Lett., 64(2017), 223–230. |
| [17] | A. Lahrouz, L. Omari and D. Kiouach, Global analysis of a deterministic and stochastic nonlinear SIRS epidemic model, Nonlinear Anal. Model. Control, 16(2011), 59–76. |
| [18] | A. Lahrouz, L. Omari and D. Kiouach, Extinction and stationary distribution of a stochastic SIRS epidemic model with non-linear incidence, Stat. Probabil. Lett., 83(2013), 960–968. |
| [19] | Q. Liu, D. Jiang, N. Shi, et al., Stationary distribution and extinction of a stochastic SIRS epidemic model with standard incidence, Physica A, 469(2017), 510–517. |
| [20] | Y.Zhao and D. Jiang, Thethreshold of a stochastic SIRS epidemic model withsaturated incidence, Appl. Math. Lett., 34(2014), 90–93. |
| [21] | Q. Liu and Q. Chen, Dynamics of a stochastic SIR epidemic model with saturated incidence, Appl. Math. Comput., 282(2016), 155–166. |
| [22] | Y. Song, A. Miao, T. Zhang, et al., Extinction and persistence of a stochastic SIRS epidemic model with saturated incidence rate and transfer from infectious to susceptible, Adv. Differ. Equ., 293(2018). |
| [23] | T. Tang, Z. Teng and Z. Li, Threshold behavior in a class of stochastic SIRS epidemic models with nonlinear incidence, Stoch. Anal. Appl., 33(2015), 994–1019. |
| [24] | D. Xiao and S. Ruan, Global analysis of an epidemic model with nonmonotone incidence rate, Math. Biosci., 208(2007), 419–429. |
| [25] | J. Cui, Y. Sun and H. Zhu, The impact of media on the control of infectious diseases, J. Dynam. Differ. Equat., 20(2008), 31–53. |
| [26] | T. Caraballo, M. EI Fatini, R. Pettersson, et al., A stochastic SIRI epidemic model with relapse and media coverage, Discrete Contin. Dyn. Syst., Ser. B, 23(2018), 3483–3501. |
| [27] | X. Mao, Stochsatic Differential Equations and Applications, Second edition, Horwood Publishing Limited, Chichester, 2007. |
| [28] | M. Liu and K. Wang, Dynamics of a Leslie-Gower Holling-type II predator-prey system with Lévy jumps, Nonlinear Anal., 85(2013), 204–213. |
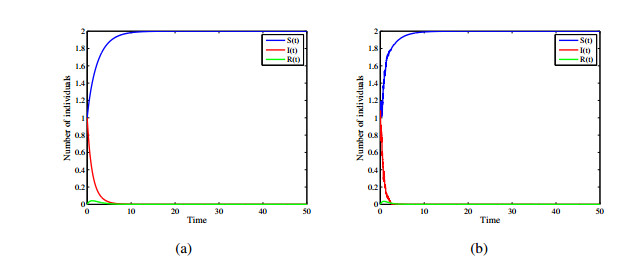
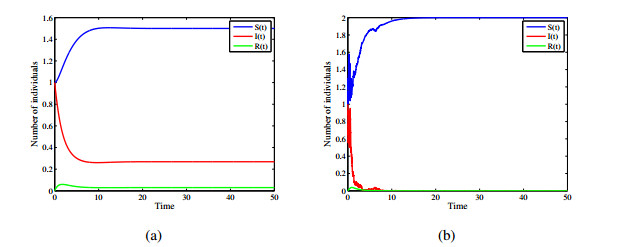
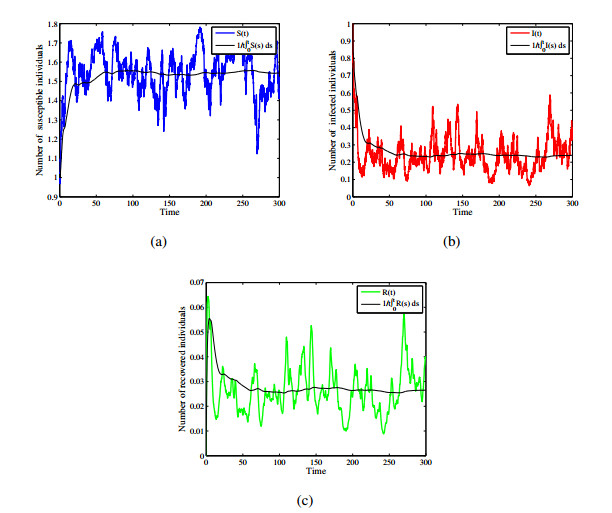
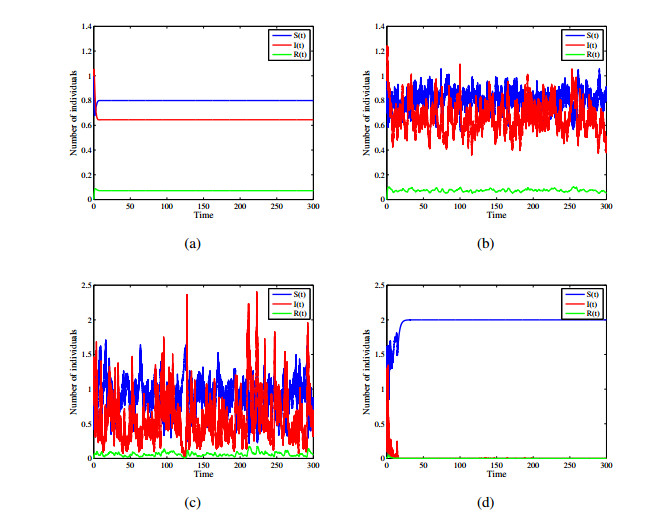
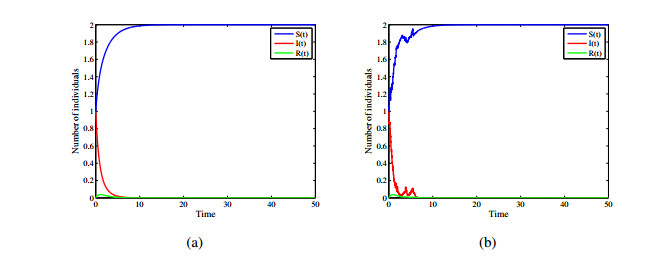
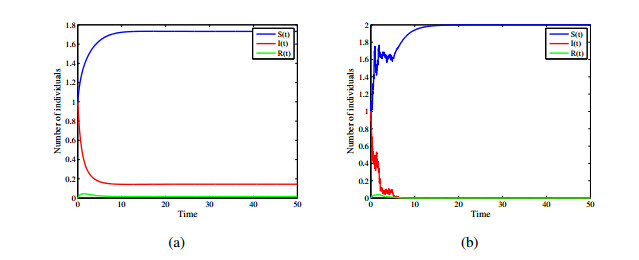
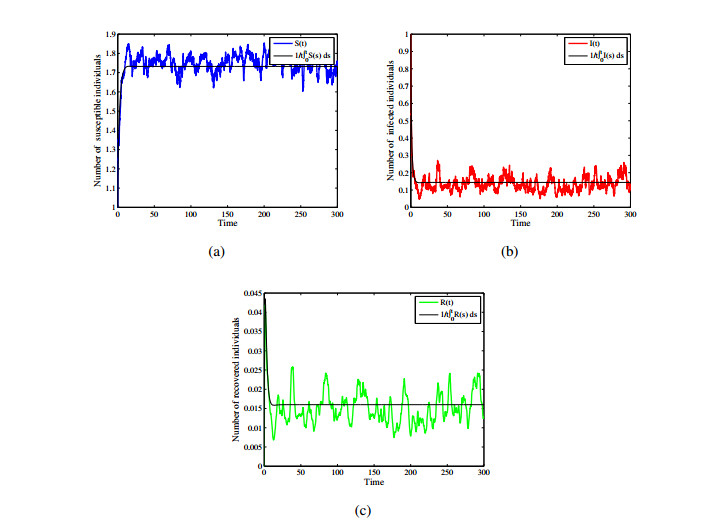
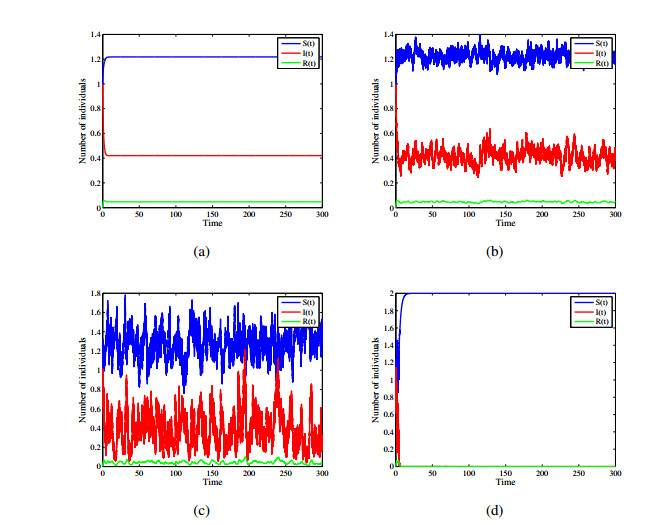
Figures(12)

Yanmei Wang, Guirong Liu. Dynamics analysis of a stochastic SIRS epidemic model with nonlinear incidence rate and transfer from infectious to susceptible[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 6047-6070. doi: 10.3934/mbe.2019303







 DownLoad:
DownLoad: