Citation: Moitri Sen, Malay Banerjee, Yasuhiro Takeuchi. Influence of Allee effect in prey populations on the dynamics of two-prey-one-predator model[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 883-904. doi: 10.3934/mbe.2018040

| [1] | [ P. Aguirre,E. González-Olivares,E. Sáez, Three limit cycles in a Leslie-Gower predator-prey model with additive Allee effect, SIAM Journal on Applied Mathematics, 69 (2009): 1244-1262. |
| [2] | [ P. Aguirre,E. González-Olivares,E. Sáez, Two limit cycles in a Leslie-Gower predator-prey model with additive Allee effect, Nonlinear Analysis: Real World Applications, 10 (2009): 1401-1416. |
| [3] | [ W. C. Allee, Animal Aggregations: A study in general sociology, The Quarterly Review of Biology, 2 (1927): 367-398. |
| [4] | [ L. Berec,E. Angulo,F. Courchamp, Multiple Allee effects and population management, Trends in Ecology & Evolution, 22 (2007): 185-191. |
| [5] | [ F. Berezovskaya,S. Wirkus,B. Song,C. Castillo-Chavez, Dynamics of population communities with prey migrations and Allee effects: a bifurcation approach, Mathematical Medicine and Biology, 28 (2011): 129-152. |
| [6] | [ E. D. Conway,J. A. Smoller, Global analysis of a system of predator-prey equations, SIAM Journal on Applied Mathematics, 46 (1986): 630-642. |
| [7] | [ F. Courchamp,T. Clutton-Brock,B. Grenfell, Inverse density dependence and the Allee effect, Trends in Ecology & Evolution, 14 (1999): 405-410. |
| [8] | [ B. Dennis, Allee effects: Population growth, critical density, and the chance of extinction, Natural Resource Modeling, 3 (1989): 481-538. |
| [9] | [ Y. C. Lai and R. L. Winslow, Geometric properties of the chaotic saddle responsible for supertransients in spatiotemporal chaotic systems Physical Review Letters, 74 (1995), p5208. |
| [10] | [ M. A. Lewis,P. Kareiva, Allee dynamics and the spread of invading organisms, Theoretical Population Biology, 43 (1993): 141-158. |
| [11] | [ A. J. Lotka, A Natural Population Norm I & Ⅱ, 1913. |
| [12] | [ A. Morozov,S. Petrovskii,B.-L. Li, Spatiotemporal complexity of patchy invasion in a predator-prey system with the Allee effect, Journal of Theoretical Biology, 238 (2006): 18-35. |
| [13] | [ A. Y. Morozov,M. Banerjee,S. V. Petrovskii, Long-term transients and complex dynamics of a stage-structured population with time delay and the Allee effect, Journal of Theoretical Biology, 396 (2016): 116-124. |
| [14] | [ M. Sen,M. Banerjee,A. Morozov, Bifurcation analysis of a ratio-dependent prey-predator model with the Allee effect, Ecological Complexity, 11 (2012): 12-27. |
| [15] | [ M. Sen,M. Banerjee, Rich global dynamics in a prey-predator model with Allee effect and density dependent death rate of predator, International Journal of Bifurcation and Chaos, 25 (2015): 17pp. |
| [16] | [ P. A. Stephens,W. J. Sutherland, Consequences of the allee effect for behaviour, ecology and conservation, Trends in Ecology & Evolution, 14 (1999): 401-405. |
| [17] | [ Y. Takeuchi, Global Dynamical Properties of Lotka-Volterra Systems, World Scientific, 1996. |
| [18] | [ Y. Takeuchi,N. Adachi, Existence and bifurcation of stable equilibrium in two-prey, one-predator communities, Bulletin of Mathematical Biology, 45 (1983): 877-900. |
| [19] | [ V. Volterra, Fluctuations in the abundance of a species considered mathematically, Nature, 118 (1926): 558-560. |
| [20] | [ G. Wang,X. G. Liang,F. Z. Wang, The competitive dynamics of populations subject to an Allee effect, Ecological Modelling, 124 (1999): 183-192. |
| [21] | [ J. Zu,M. Mimura, The impact of Allee effect on a predator-prey system with Holling type ii functional response, Applied Mathematics and Computation, 217 (2010): 3542-3556. |
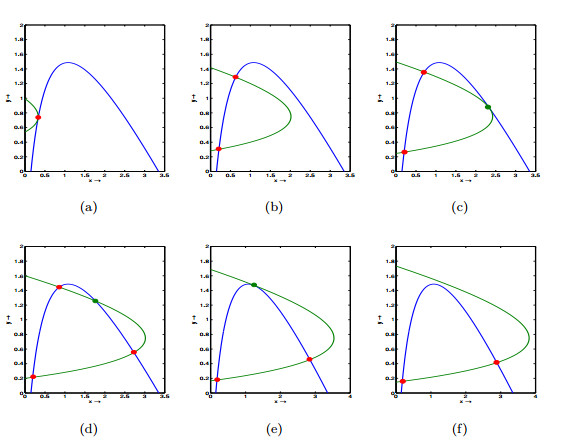
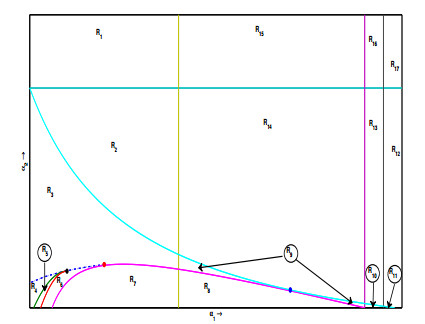
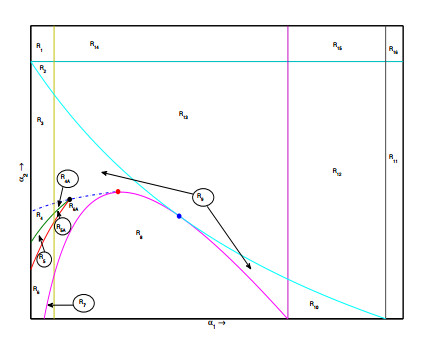
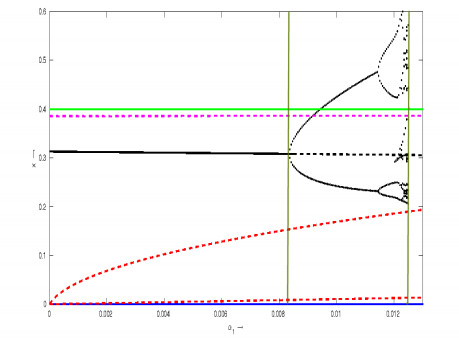
Figures(6) / Tables(3)

Moitri Sen, Malay Banerjee, Yasuhiro Takeuchi. Influence of Allee effect in prey populations on the dynamics of two-prey-one-predator model[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 883-904. doi: 10.3934/mbe.2018040







 DownLoad:
DownLoad: