Citation: Claudio Arancibia–Ibarra, José Flores. Modelling and analysis of a modified May-Holling-Tanner predator-prey model with Allee effect in the prey and an alternative food source for the predator[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8052-8073. doi: 10.3934/mbe.2020408

| [1] |
I. Hanski, L. Hansson, H. Henttonen, Specialist predators, generalist predators, and the microtine rodent cycle, J. Anim. Ecol., 60 (1991), 353-367. doi: 10.2307/5465

|
| [2] | I. Hanski, H. Henttonen, E. Korpimäki, L. Oksanen, P. Turchin, Small-rodent dynamics and predation, Ecology, 82 (2001), 15005-1520. |
| [3] |
P. Turchin, I. Hanski, An empirically based model for latitudinal gradient in vole population dynamics, Am. Nat., 149 (1997), 842-874. doi: 10.1086/286027
|
| [4] | T. Dalkvist, R. Sibly, C. Topping, How predation and landscape fragmentation affect vole population dynamics, Plos One, 149 (2011), 1-8. |
| [5] |
D. Madison, R. FitzGerald, W. McShea, Dynamics of social nesting in overwintering meadow voles (Microtus pennsylvanicus): possible consequences for population cycling, Behav. Ecol. Sociobiol., 15 (1984), 9-17. doi: 10.1007/BF00310209
|
| [6] |
M. Graham, X. Lambin, The impact of weasel predation on cyclic field-vole survival: the specialist predator hypothesis contradicted, J. Anim. Ecol., 71 (2002), 946-956. doi: 10.1046/j.1365-2656.2002.00657.x
|
| [7] | P. Turchin, Complex population dynamics: a theoretical/empirical synthesis, Monographs in population biology, Princeton University Press, Princeton N.J., 2003. |
| [8] | R. May, Stability and complexity in model ecosystems, Monographs in population biology, Princeton University Press, Princeton N.J., 1974. |
| [9] |
p. Leslie, J. Gower, The Properties of a Stochastic Model for the Predator-Prey Type of Interaction Between Two Species, Biometrika, 47 (1960), 219-234. doi: 10.1093/biomet/47.3-4.219
|
| [10] | E. Pielou, An introduction to Mathematical Ecology, Wiley-Interscience, New York, 1974. |
| [11] |
X. Santos, M. Cheylan, Taxonomic and functional response of a Mediterranean reptile assemblage to a repeated fire regime, Biol. Conserv., 168 (2013), 90-98. doi: 10.1016/j.biocon.2013.09.008
|
| [12] | P. Aguirre, E. González-Olivares, E. Sáez, Two limit cycles in a Leslie-Gower predator-prey model with additive Allee effect, Nonlinear Anal.-Real., 10 (2009), 1401-1416. |
| [13] |
C. Arancibia-Ibarra, The basins of attraction in a Modified May-Holling-Tanner predator-prey model with Allee effect, Nonlinear Anal.-Theor., 185 (2019), 15-28. doi: 10.1016/j.na.2019.03.004
|
| [14] | E. Sáez, E. González-Olivares, Dynamics on a predator-prey model, SIAM J. Appl. Math., 59 (1999), 1867-1878. |
| [15] | F. Dumortier, J. Llibre, J. C. Artés, Qualitative theory of planar differential systems, Springer Berlin Heidelberg, Springer-Verlag Berlin Heidelberg, 2006. |
| [16] |
B. Lisena, Global stability of a periodic Holling-Tanner predator-prey model, Math. Method. Appl. Sci., 41 (2018), 3270-3281. doi: 10.1002/mma.4814
|
| [17] |
H. Cao, Z. Yue, Y. Zhou, The stability and bifurcation analysis of a discrete Holling-Tanner model, Adv. Differ. Equ., 2013 (2013), 1-17. doi: 10.1186/1687-1847-2013-1
|
| [18] | W. Allee, The social life of animals, WW Norton & Co, New York, 1938. |
| [19] | W. Allee, O. Park, A. Emerson, T. Park, K. Schmidt, Principles of animal ecology, WB Saundere Co. Ltd., Philadelphia, 1949. |
| [20] |
L. Berec, E. Angulo, F. Courchamp, Multiple Allee effects and population management, Trends. Ecol. Evol., 22 (2007), 185-191. doi: 10.1016/j.tree.2006.12.002
|
| [21] |
G. Buffoni, M. Groppi, C. Soresina, Dynamics of predator-prey models with a strong Allee effect on the prey and predator-dependent trophic functions, Nonlinear Anal.-Real., 30 (2016), 143-169. doi: 10.1016/j.nonrwa.2015.12.001
|
| [22] |
G. van Voorn, L. Hemerik, M. Boer, B. Kooi, Heteroclinic orbits indicate overexploitation in predator-prey systems with a strong Allee effect, Math. Biosci., 209 (2007), 451-469. doi: 10.1016/j.mbs.2007.02.006
|
| [23] |
J. Wang, J. Shi, J. Wei, Predator-prey system with strong Allee effect in prey, J. Math. Biol., 62 (2011), 291-331. doi: 10.1007/s00285-010-0332-1
|
| [24] |
article P. Stephens, W. Sutherland, R. Freckleton, What is the Allee effect?, Oikos, 87 (1999), 185-190. doi: 10.2307/3547011
|
| [25] |
M. Jankovic, S. Petrovskii, Are time delays always destabilizing? Revisiting the role of time delays and the Allee effect, Theor. Ecol., 7 (2014), 335-349. doi: 10.1007/s12080-014-0222-z
|
| [26] |
D. Morris, Measuring the Allee effect: positive density dependence in small mammals, Ecology, 83 (2002), 14-20. doi: 10.1890/0012-9658(2002)083[0014:MTAEPD]2.0.CO;2
|
| [27] |
R. Ostfeld, C. Canham, Density-dependent processes in meadow voles: an experimental approach, Ecology, 76 (1995), 521-532. doi: 10.2307/1941210
|
| [28] | F. Courchamp, L. Berec, J. Gascoigne, Allee effects in ecology and conservation, Oxford University Press, 2008. |
| [29] |
M. Liermann, R. Hilborn, Depensation: evidence, models and implications, Fish. Fish., 2 (2001), 33-58, doi: 10.1046/j.1467-2979.2001.00029.x
|
| [30] |
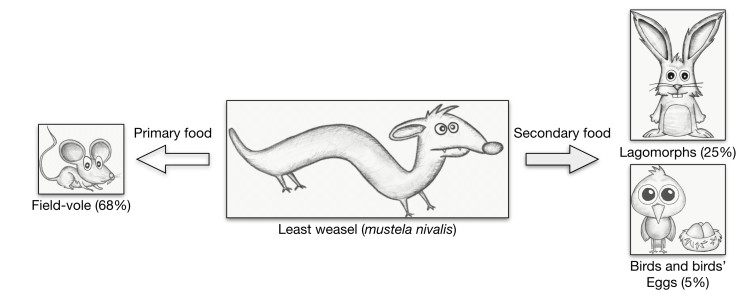
R. McDonald, C. Webbon, S. Harris, The diet of stoats (Mustela erminea) and weasels (Mustela nivalis) in Great Britain, J. Zool., 252 (2000), 363-371. doi: 10.1111/j.1469-7998.2000.tb00631.x
|
| [31] |
M. Andersson, S. Erlinge, Influence of predation on rodent populations, Oikos, 29 (1977), 591-597. doi: 10.2307/3543597
|
| [32] |
S. Erlinge, Predation and noncyclicity in a microtine population in southern Sweden, Oikos, 50 (1987), 347-352. doi: 10.2307/3565495
|
| [33] |
I. Hanski, P. Turchin, E. Korpimaki, H. Henttonen, Population oscillations of boreal rodents: regulation by mustelid predators leads to chaos, Nature, 364 (1993), 232-235. doi: 10.1038/364232a0
|
| [34] | L. Hansson, Competition between rodents in successional stages of taiga forests: Microtus agrestis vs. Clethrionomys glareolus, Oikos, 40 (1983), 258-266. |
| [35] |
D. Wollkind, J. Collings, J. Logan, Metastability in a temperature-dependent model system for predator-prey mite outbreak interactions on fruit trees, B. Math. Biol., 50 (1988), 379-409. doi: 10.1016/S0092-8240(88)90005-5
|
| [36] |
M. Aziz-Alaoui, M. Daher, Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type II schemes, Appl. Math. Lett., 16 (2003), 1069-1075. doi: 10.1016/S0893-9659(03)90096-6
|
| [37] |
A. Dhooge, W. Govaerts, V. Kuznetsov, H. Meijer, B. Sautois, New features of the software MatCont for bifurcation analysis of dynamical systems, Math. Comp. Model. Dyn., 14 (2008), 147-175. doi: 10.1080/13873950701742754
|
| [38] | J. Huang, Y. Gong, S. Ruan, Bifurcation analysis in a predator-prey model with constant-yield predator harvesting, Discrete. Cont. Dyn-B., 18 (2013), 2101-2121. |
| [39] |
P. Roux, J. Shaw, S. Chown, Ontogenetic shifts in plant interactions vary with environmental severity and affect population structure, New. Phytol., 200 (2013), 241-250. doi: 10.1111/nph.12349
|
| [40] | C. Arancibia-Ibarra, J. Flores, G. Pettet, P. van Heijster, A Holling-Tanner predator-prey model with strong Allee effect, Int. J. Bifurcat. Chaos, 29 (2019), 1-16. |
| [41] | C. Arancibia-Ibarra, E. González-Olivares, The Holling-Tanner model considering an alternative food for predator, Proc. CMMSE 2015, (2015), 130-141. |
| [42] | E. González-Olivares, C. Arancibia-Ibarra, A. Rojas-Palma, B. González-Ya?ez, Bifurcations and multistability on the May-Holling-Tanner predation model considering alternative food for the predators, Math. Biosci. Eng., 16 (2019), 4274-4298. |
| [43] | C. Arancibia-Ibarra, E. González-Olivares, A modified Leslie-Gower predator-prey model with hyperbolic functional response and Allee effect on prey, BIOMAT Int. S. Math. Co., 1(2011), 46-162. |
| [44] | E. González-Olivares, C. Arancibia-Ibarra, A. Rojas-Palma, B. González-Ya?ez, Dynamics of a modified Leslie-Gower predation model considering a generalist predator and the hyperbolic functional response, Math. Biosci. Eng., 16 (2019), 7995-8024. |
Figures(7) / Tables(2)

Claudio Arancibia–Ibarra, José Flores. Modelling and analysis of a modified May-Holling-Tanner predator-prey model with Allee effect in the prey and an alternative food source for the predator[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8052-8073. doi: 10.3934/mbe.2020408







 DownLoad:
DownLoad: