Autophagy is a normal cellular physiological process. As one of the cell degradation systems, it participates in the lysosomal pathway process of degrading damaged proteins and subcellular organelles to maintain cell metabolism and energy states. Moreover, autophagy is essential for regulating organelle quality control and cell homeostasis. In recent decades, a large number of studies have demonstrated that autophagy abnormalities are present in a variety of human malignancies and that autophagy plays a crucial role in all stages of tumor development. Multiple tumors interfere with autophagy's normal regulation and use autophagy's essential properties to restructure their proteome, reprogram their metabolism, and adapt to stress. This article primarily discusses how autophagy either promotes or inhibits cancer development, the significance of autophagy in maintaining cell genome stability, and the role of selective autophagy in the reshaping and quality control of tumor cells.
Citation: Chunzheng Li, Chenyu Wei, Gongke Zhao, Xianguang Yang. Cancer cells remodeling and quality control are inextricably linked to autophagy[J]. AIMS Molecular Science, 2023, 10(2): 109-126. doi: 10.3934/molsci.2023009
Autophagy is a normal cellular physiological process. As one of the cell degradation systems, it participates in the lysosomal pathway process of degrading damaged proteins and subcellular organelles to maintain cell metabolism and energy states. Moreover, autophagy is essential for regulating organelle quality control and cell homeostasis. In recent decades, a large number of studies have demonstrated that autophagy abnormalities are present in a variety of human malignancies and that autophagy plays a crucial role in all stages of tumor development. Multiple tumors interfere with autophagy's normal regulation and use autophagy's essential properties to restructure their proteome, reprogram their metabolism, and adapt to stress. This article primarily discusses how autophagy either promotes or inhibits cancer development, the significance of autophagy in maintaining cell genome stability, and the role of selective autophagy in the reshaping and quality control of tumor cells.
phosphatidylinositol-3-phosphate
WD-repeat protein interacting with phosphoinositides
double FYVE-containing protein 1
genetically engineered mouse models
reactive oxygen species
damage-regulated autophagy modulator 1
cancer stem cell

| [1] | Poillet-Perez L, White E (2019) Role of tumor and host autophagy in cancer metabolism. Genes Dev 33: 610-619. https://doi.org/10.1101/gad.325514.119 |
| [2] | Kaushik S, Cuervo AM (2018) The coming of age of chaperone-mediated autophagy. Nat Rev Mol Cell Biol 19: 365-381. https://doi.org/10.1038/s41580-018-0001-6 |
| [3] | Oku M, Sakai Y (2018) Three distinct types of microautophagy based on membrane dynamics and molecular machineries. Bioessays 40: 1800008. https://doi.org/10.1002/bies.201800008 |
| [4] | Yorimitsu T, Klionsky DJ (2005) Autophagy: Molecular machinery for self-eating. Cell Death Differ 12: 1542-1552. https://doi.org/10.1038/sj.cdd.4401765 |
| [5] | Anding AL, Baehrecke EH (2017) Cleaning house: Selective autophagy of organelles. Dev Cell 41: 10-22. https://doi.org/10.1016/j.devcel.2017.02.016 |
| [6] | Itakura E, Kishi C, Inoue K, et al. (2008) Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG. Mol Biol Cell 19: 5360-5372. https://doi.org/10.1091/mbc.e08-01-0080 |
| [7] | Zheng K, He Z, Kitazato K, et al. (2019) Selective autophagy regulates cell cycle in cancer therapy. Theranostics 9: 104-125. https://doi.org/10.7150/thno.30308 |
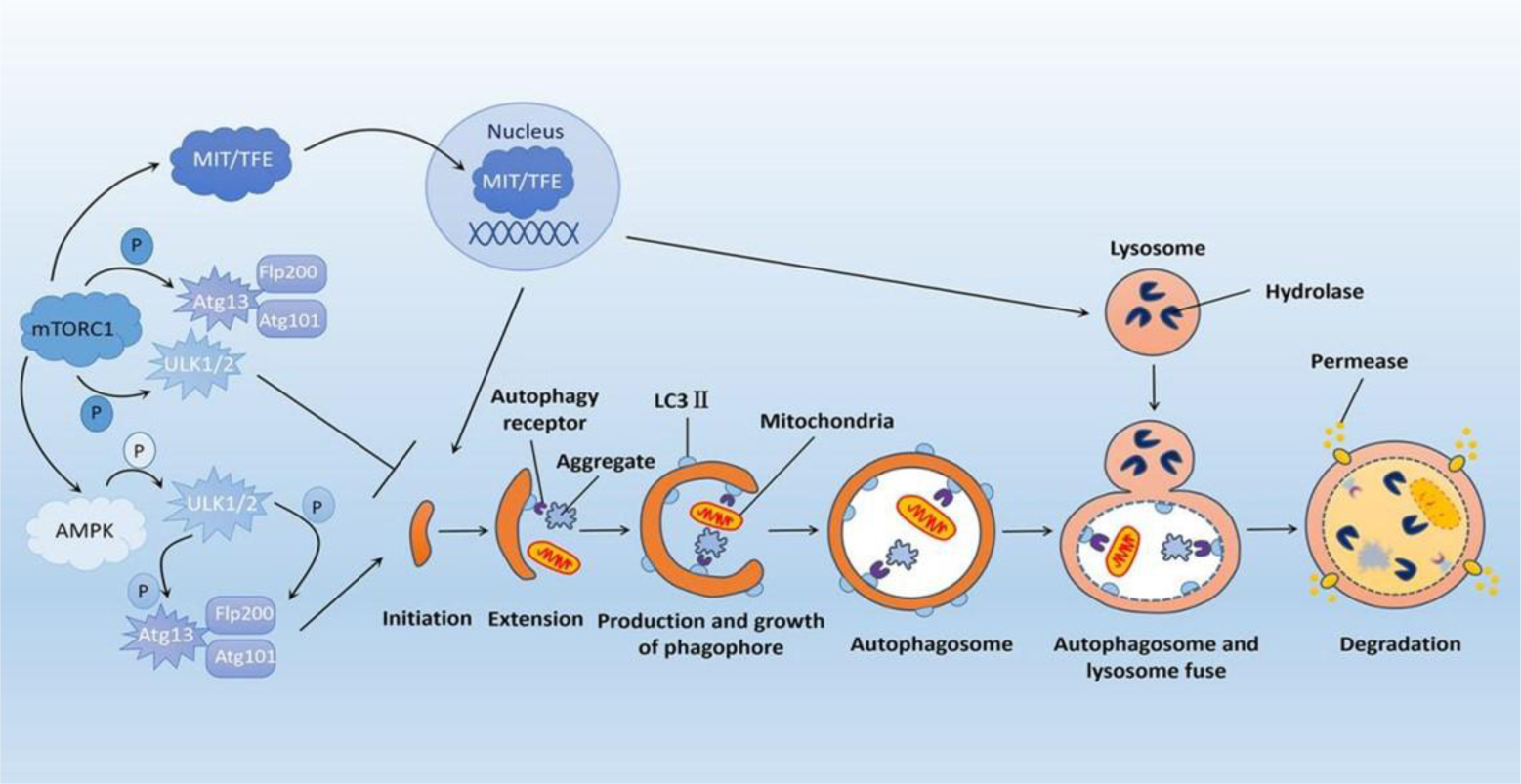
| [8] | Mizushima N (2010) The role of the Atg1/ULK1 complex in autophagy regulation. Curr Opin Cell Biol 22: 132-139. https://doi.org/10.1016/j.ceb.2009.12.004 |
| [9] | Papinski D, Kraft C (2016) Regulation of autophagy by signaling through the Atg1/ULK1 complex. J Mol Biol 428: 1725-1741. https://doi.org/10.1016/j.jmb.2016.03.030 |
| [10] | Hosokawa N, Hara T, Kaizuka T, et al. (2009) Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy. Mol Biol Cell 20: 1981-1991. https://doi.org/10.1091/mbc.e08-12-1248 |
| [11] | Xu F, Na L, Li Y, et al. (2020) Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci 10: 54. https://doi.org/10.1186/s13578-020-00416-0 |
| [12] | Mizushima N, Noda T, Ohsumi Y (1999) Apg16p is required for the function of the Apg12p-Apg5p conjugate in the yeast autophagy pathway. EMBO J 18: 3888-3896. https://doi.org/10.1093/emboj/18.14.3888 |
| [13] | Xie Z, Klionsky DJ (2007) Autophagosome formation: Core machinery and adaptations. Nat Cell Biol 9: 1102-1109. https://doi.org/10.1038/ncb1007-1102 |
| [14] | Sakoh-Nakatogawa M, Matoba K, Asai E, et al. (2013) Atg12-Atg5 conjugate enhances E2 activity of Atg3 by rearranging its catalytic site. Nat Struct Mol Biol 20: 433-439. https://doi.org/10.1038/nsmb.2527 |
| [15] | Ichimura Y, Kirisako T, Takao T, et al. (2000) A ubiquitin-like system mediates protein lipidation. Nature 408: 488-492. https://doi.org/10.1038/35044114 |
| [16] | Kabeya Y, Mizushima N, Ueno T, et al. (2000) LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J 19: 5720-5728. https://doi.org/10.1093/emboj/19.21.5720 |
| [17] | Mizushima N, Klionsky DJ (2007) Protein turnover via autophagy: Implications for metabolism. Annu Rev Nutr 27: 19-40. https://doi.org/10.1146/annurev.nutr.27.061406.093749 |
| [18] | Tan Q, Wang M, Yu M, et al. (2016) Role of autophagy as a survival mechanism for hypoxic cells in tumors. Neoplasia 18: 347-355. https://doi.org/10.1016/j.neo.2016.04.003 |
| [19] | B'Chir W, Maurin AC, Carraro V, et al. (2013) The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res 41: 7683-7699. https://doi.org/10.1093/nar/gkt563 |
| [20] | Aita VM, Liang XH, Murty VV, et al. (1999) Cloning and genomic organization of beclin 1, a candidate tumor suppressor gene on chromosome 17q21. Genomics 59: 59-65. https://doi.org/10.1006/geno.1999.5851 |
| [21] | Liang XH, Jackson S, Seaman M, et al. (1999) Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature 402: 672-676. https://doi.org/10.1038/45257 |
| [22] | Butera G, Mullappilly N, Masetto F, et al. (2019) Regulation of autophagy by nuclear GAPDH and its aggregates in cancer and neurodegenerative disorders. Int J Mol Sci 20: 2062. https://doi.org/10.3390/ijms20092062 |
| [23] | An CH, Kim MS, Yoo NJ, et al. (2011) Mutational and expressional analyses of ATG5, an autophagy-related gene, in gastrointestinal cancers. Pathol Res Pract 207: 433-437. https://doi.org/10.1016/j.prp.2011.05.002 |
| [24] | Inami Y, Waguri S, Sakamoto A, et al. (2011) Persistent activation of Nrf2 through p62 in hepatocellular carcinoma cells. J Cell Biol 193: 275-284. https://doi.org/10.1083/jcb.201102031 |
| [25] | Mariño G, Salvador-Montoliu N, Fueyo A, et al. (2007) Tissue-specific autophagy alterations and increased tumorigenesis in mice deficient in Atg4C/autophagin-3. J Biol Chem 282: 18573-18583. https://doi.org/10.1074/jbc.M701194200 |
| [26] | Mathew R, Karp CM, Beaudoin B, et al. (2009) Autophagy suppresses tumorigenesis through elimination of p62. Cell 137: 1062-1075. https://doi.org/10.1016/j.cell.2009.03.048 |
| [27] | Yang A, Herter-Sprie G, Zhang H, et al. (2018) Autophagy sustains pancreatic cancer growth through both cell-autonomous and nonautonomous mechanisms. Cancer Discov 8: 276-287. https://doi.org/10.1158/2159-8290.CD-17-0952 |
| [28] | Choudhury S, Kolukula V, Preet A, et al. (2013) Dissecting the pathways that destabilize mutant p53: The proteasome or autophagy?. Cell Cycle 12: 1022-1029. https://doi.org/10.4161/cc.24128 |
| [29] | Athar M, Elmets CA, Kopelovich L (2011) Pharmacological activation of p53 in cancer cells. Curr Pharm Des 17: 631-639. https://doi.org/10.2174/138161211795222595 |
| [30] | Feng Z, Zhang H, Levine AJ, et al. (2005) The coordinate regulation of the p53 and mTOR pathways in cells. Proc Natl Acad Sci USA 102: 8204-8209. https://doi.org/10.1073/pnas.0502857102 |
| [31] | Crighton D, Wilkinson S, O'Prey J, et al. (2006) DRAM, a p53-induced modulator of autophagy, is critical for apoptosis. Cell 126: 121-134. https://doi.org/10.1016/j.cell.2006.05.034 |
| [32] | Guo JY, Chen HY, Mathew R, et al. (2011) Activated Ras requires autophagy to maintain oxidative metabolism and tumorigenesis. Genes Dev 25: 460-470. https://doi.org/10.1101/gad.2016311 |
| [33] | Koustas E, Sarantis P, Papavassiliou AG, et al. (2018) Upgraded role of autophagy in colorectal carcinomas. World J Gastrointest Oncol 10: 367-369. https://doi.org/10.4251/wjgo.v10.i11.367 |
| [34] | Alves S, Castro L, Fernandes MS, et al. (2015) Colorectal cancer-related mutant KRAS alleles function as positive regulators of autophagy. Oncotarget 6: 30787-30802. http://doi.org/10.18632/oncotarget.5021 |
| [35] | Kahraman DC, Kahraman T, Cetin-Atalay R (2019) Targeting PI3K/Akt/mTOR pathway identifies differential expression and functional role of IL8 in liver cancer stem cell enrichment. Mol Cancer Ther 18: 2146-2157. https://doi.org/10.1158/1535-7163.Mct-19-0004 |
| [36] | Kim TW, Lee SJ, Kim JT, et al. (2016) Kallikrein-related peptidase 6 induces chemotherapeutic resistance by attenuating auranofin-induced cell death through activation of autophagy in gastric cancer. Oncotarget 7: 85332-85348. https://doi.org/10.18632/oncotarget.13352 |
| [37] | Xin L, Zhou Q, Yuan YW, et al. (2019) METase/lncRNA HULC/FoxM1 reduced cisplatin resistance in gastric cancer by suppressing autophagy. J Cancer Res Clin Oncol 145: 2507-2517. https://doi.org/10.1007/s00432-019-03015-w |
| [38] | Tschan MP, Simon HU (2010) The role of autophagy in anticancer therapy: promises and uncertainties. J Intern Med 268: 410-418. https://doi.org/10.1111/j.1365-2796.2010.02266.x |
| [39] | Vessoni AT, Filippi-Chiela EC, Menck CF, et al. (2013) Autophagy and genomic integrity. Cell Death Differ 20: 1444-1454. https://doi.org/10.1038/cdd.2013.103 |
| [40] | Hewitt G, Korolchuk VI (2017) Repair, reuse, recycle: The expanding role of autophagy in genome maintenance. Trends Cell Biol 27: 340-351. https://doi.org/10.1016/j.tcb.2016.11.011 |
| [41] | Chen H, Wang J, Liu Z, et al. (2016) Mitochondrial DNA depletion causes decreased ROS production and resistance to apoptosis. Int J Mol Med 38: 1039-1046. https://doi.org/10.3892/ijmm.2016.2697 |
| [42] | Chao T, Shih HT, Hsu SC, et al. (2021) Autophagy restricts mitochondrial DNA damage-induced release of ENDOG (endonuclease G) to regulate genome stability. Autophagy 17: 3444-3460. https://doi.org/10.1080/15548627.2021.1874209 |
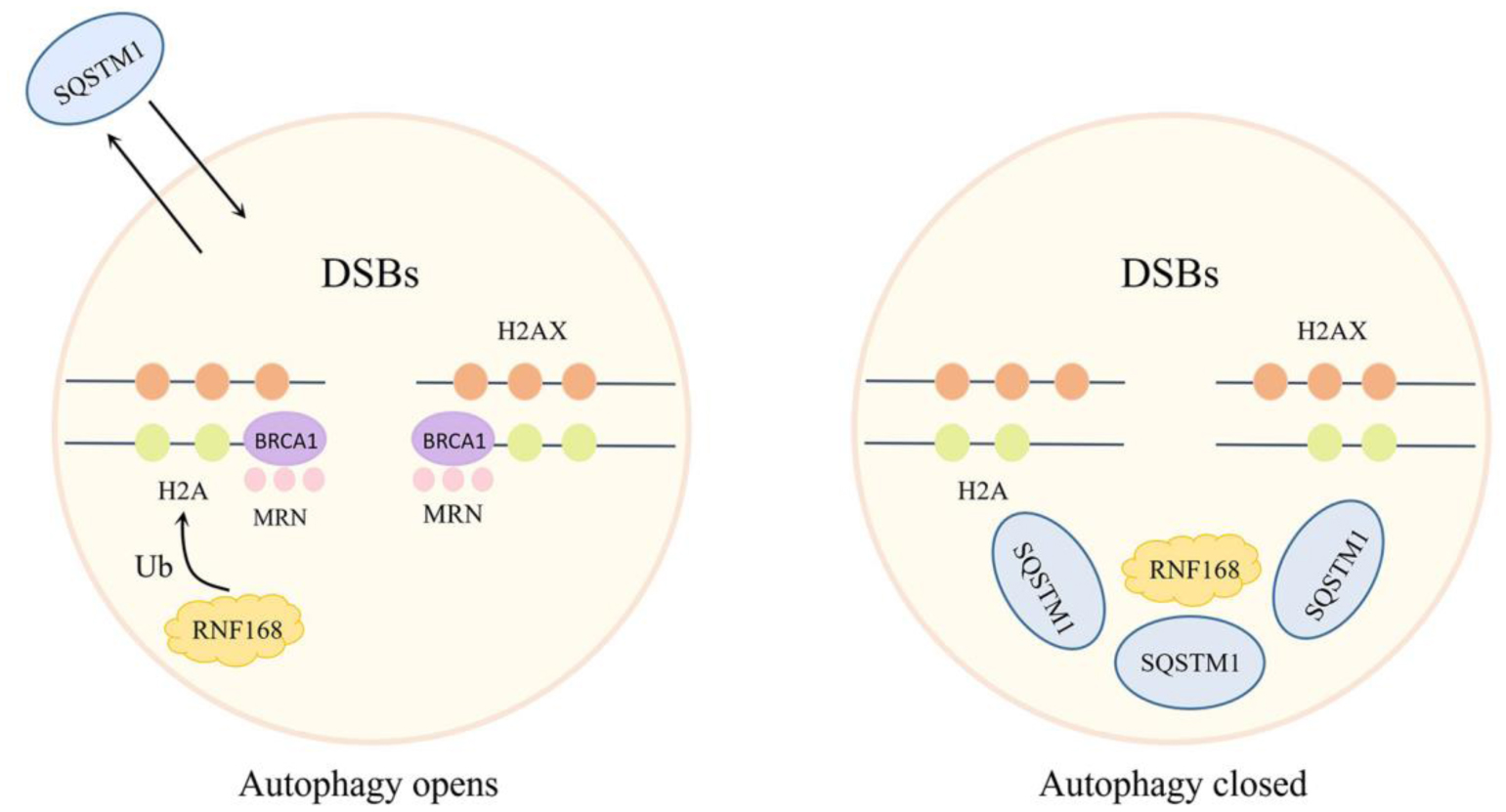
| [43] | Robert T, Vanoli F, Chiolo I, et al. (2011) HDACs link the DNA damage response, processing of double-strand breaks and autophagy. Nature 471: 74-79. https://doi.org/10.1038/nature09803 |
| [44] | Belaid A, Cerezo M, Chargui A, et al. (2013) Autophagy plays a critical role in the degradation of active RHOA, the control of cell cytokinesis, and genomic stability. Cancer Res 73: 4311-4322. |
| [45] | Youle RJ, Narendra DP (2011) Mechanisms of mitophagy. Nat Rev Mol Cell Biol 12: 9-14. |
| [46] | Ryter SW, Bhatia D, Choi ME (2019) Autophagy: A lysosome-dependent process with implications in cellular redox homeostasis and human disease. Antioxid Redox Signal 30: 138-159. |
| [47] | Yao RQ, Ren C, Xia ZF, et al. (2021) Organelle-specific autophagy in inflammatory diseases: A potential therapeutic target underlying the quality control of multiple organelles. Autophagy 17: 385-401. https://doi.org/10.1080/15548627.2020.1725377 |
| [48] | Springer MZ, Macleod KF (2016) In brief: Mitophagy: Mechanisms and role in human disease. J Pathol 240: 253-255. https://doi.org/10.1002/path.4774 |
| [49] | Strappazzon F, Nazio F, Corrado M, et al. (2015) AMBRA1 is able to induce mitophagy via LC3 binding, regardless of PARKIN and p62/SQSTM1. Cell Death Differ 22: 419-432. https://doi.org/10.1038/cdd.2014.139 |
| [50] | Papadopoulos C, Kravic B, Meyer H (2020) Repair or lysophagy: Dealing with damaged lysosomes. J Mol Biol 432: 231-239. https://doi.org/10.1016/j.jmb.2019.08.010 |
| [51] | Maejima I, Takahashi A, Omori H, et al. (2013) Autophagy sequesters damaged lysosomes to control lysosomal biogenesis and kidney injury. EMBO J 32: 2336-2347. https://doi.org/10.1038/emboj.2013.171 |
| [52] | Fraiberg M, Elazar Z (2016) A TRIM16-Galactin3 complex mediates autophagy of damaged endomembranes. Dev Cell 39: 1-2. https://doi.org/10.1016/j.devcel.2016.09.025 |
| [53] | Koerver L, Papadopoulos C, Liu B, et al. (2019) The ubiquitin-conjugating enzyme UBE2QL1 coordinates lysophagy in response to endolysosomal damage. EMBO Rep 20: e48014. https://doi.org/10.15252/embr.201948014 |
| [54] | Papadopoulos C, Kirchner P, Bug M, et al. (2017) VCP/p97 cooperates with YOD1, UBXD1 and PLAA to drive clearance of ruptured lysosomes by autophagy. EMBO J 36: 135-150. https://doi.org/10.15252/embj.201695148 |
| [55] | Lu Z, Miao Y, Muhammad I, et al. (2017) Colistin-induced autophagy and apoptosis involves the JNK-Bcl2-Bax signaling pathway and JNK-p53-ROS positive feedback loop in PC-12 cells. Chem Biol Interact 277: 62-73. https://doi.org/10.1016/j.cbi.2017.08.011 |
| [56] | Chourasia AH, Tracy K, Frankenberger C, et al. (2015) Mitophagy defects arising from BNip3 loss promote mammary tumor progression to metastasis. EMBO Rep 16: 1145-1163. https://doi.org/10.15252/embr.201540759 |
| [57] | Sun L, Li T, Wei Q, et al. (2014) Upregulation of BNIP3 mediated by ERK/HIF-1α pathway induces autophagy and contributes to anoikis resistance of hepatocellular carcinoma cells. Future Oncol 10: 1387-1398. https://doi.org/10.2217/fon.14.70 |
| [58] | Liu K, Lee J, Kim JY, et al. (2017) Mitophagy controls the activities of tumor suppressor p53 to regulate hepatic cancer stem cells. Mol Cell 68: 281-292. https://doi.org/10.1016/j.molcel.2017.09.022 |
| [59] | Shida M, Kitajima Y, Nakamura J, et al. (2016) Impaired mitophagy activates mtROS/HIF-1α interplay and increases cancer aggressiveness in gastric cancer cells under hypoxia. Int J Oncol 48: 1379-1390. https://doi.org/10.3892/ijo.2016.3359 |
| [60] | Xie Q, Wu Q, Horbinski CM, et al. (2015) Mitochondrial control by DRP1 in brain tumor initiating cells. Nat Neurosci 18: 501-510. https://doi.org/10.1038/nn.3960 |
| [61] | Pei S, Minhajuddin M, Adane B, et al. (2018) AMPK/FIS1-Mediated mitophagy is required for self-renewal of human AML stem cells. Cell Stem Cell 23: 86-100. https://doi.org/10.1016/j.stem.2018.05.021 |
| [62] | Herzig S, Shaw RJ (2018) AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat Rev Mol Cell Biol 19: 121-135. https://doi.org/10.1038/nrm.2017.95 |
| [63] | Zuiderweg ERP, Hightower LE, Gestwicki JE (2017) The remarkable multivalency of the Hsp70 chaperones. Cell Stress Chaperones 22: 173-189. https://doi.org/10.1007/s12192-017-0776-y |
| [64] | Jego G, Hazoumé A, Seigneuric R, et al. (2013) Targeting heat shock proteins in cancer. Cancer Lett 332: 275-285. https://doi.org/10.1016/j.canlet.2010.10.014 |
| [65] | Singh KK, Ayyasamy V, Owens KM, et al. (2009) Mutations in mitochondrial DNA polymerase-γ promote breast tumorigenesis. J Hum Genet 54: 516-524. https://doi.org/10.1038/jhg.2009.71 |
| [66] | Singh B, Owens KM, Bajpai P, et al. (2015) Mitochondrial DNA polymerase POLG1 disease mutations and germline variants promote tumorigenic properties. PLoS One 10: e0139846. https://doi.org/10.1371/journal.pone.0139846 |
| [67] | MacVicar T, Langer T (2016) OPA1 processing in cell death and disease-the long and short of it. J Cell Sci 129: 2297-2306. https://doi.org/10.1242/jcs.159186 |
| [68] | Alavi MV (2019) Targeted OMA1 therapies for cancer. Int J Cancer 145: 2330-2341. https://doi.org/10.1002/ijc.32177 |
| [69] | Esteban-Martínez L, Sierra-Filardi E, McGreal RS, et al. (2017) Programmed mitophagy is essential for the glycolytic switch during cell differentiation. EMBO J 36: 1688-1706. https://doi.org/10.15252/embj.201695916 |
| [70] | Desideri E, Vegliante R, Cardaci S, et al. (2014) MAPK14/p38α-dependent modulation of glucose metabolism affects ROS levels and autophagy during starvation. Autophagy 10: 1652-1665. https://doi.org/10.4161/auto.29456 |
| [71] | Tan VP, Miyamoto S (2015) HK2/hexokinase-II integrates glycolysis and autophagy to confer cellular protection. Autophagy 11: 963-964. https://doi.org/10.1080/15548627.2015.1042195 |
| [72] | Yang X, Cheng Y, Li P, et al. (2015) A lentiviral sponge for miRNA-21 diminishes aerobic glycolysis in bladder cancer T24 cells via the PTEN/PI3K/AKT/mTOR axis. Tumour Biol 36: 383-391. https://doi.org/10.1007/s13277-014-2617-2 |
| [73] | Jiao L, Zhang HL, Li DD, et al. (2018) Regulation of glycolytic metabolism by autophagy in liver cancer involves selective autophagic degradation of HK2 (hexokinase 2). Autophagy 14: 671-684. https://doi.org/10.1080/15548627.2017.1381804 |
| [74] | Fan Q, Yang L, Zhang X, et al. (2018) Autophagy promotes metastasis and glycolysis by upregulating MCT1 expression and Wnt/β-catenin signaling pathway activation in hepatocellular carcinoma cells. J Exp Clin Cancer Res 37: 9. https://doi.org/10.1186/s13046-018-0673-y |
| [75] | de Cedrón MG, de Molina AR (2016) Microtargeting cancer metabolism: Opening new therapeutic windows based on lipid metabolism. J Lipid Res 57: 193-206. https://doi.org/10.1194/jlr.R061812 |
| [76] | Corbet C, Feron O (2017) Emerging roles of lipid metabolism in cancer progression. Curr Opin Clin Nutr Metab Care 20: 254-260. https://doi.org/10.1097/mco.0000000000000381 |
| [77] | Schlaepfer IR, Rider L, Rodrigues LU, et al. (2014) Lipid catabolism via CPT1 as a therapeutic target for prostate cancer. Mol Cancer Ther 13: 2361-2371. https://doi.org/10.1158/1535-7163.Mct-14-0183 |
| [78] | Tabe Y, Yamamoto S, Saitoh K, et al. (2017) Bone marrow adipocytes facilitate fatty acid oxidation activating AMPK and a transcriptional network supporting survival of acute monocytic leukemia cells. Cancer Res 77: 1453-1464. https://doi.org/10.1158/0008-5472.Can-16-1645 |
| [79] | Chen Y, Li X, Guo L, et al. (2015) Combining radiation with autophagy inhibition enhances suppression of tumor growth and angiogenesis in esophageal cancer. Mol Med Rep 12: 1645-1652. https://doi.org/10.3892/mmr.2015.3623 |
| [80] | Sousa CM, Biancur DE, Wang X, et al. (2016) Pancreatic stellate cells support tumour metabolism through autophagic alanine secretion. Nature 536: 479-483. https://doi.org/10.1038/nature19084 |
| [81] | Höhr AIC, Lindau C, Wirth C, et al. (2018) Membrane protein insertion through a mitochondrial β-barrel gate. Science 359. https://doi.org/10.1126/science.aah6834 |
| [82] | Christian BE, Spremulli LL (2010) Preferential selection of the 5′-terminal start codon on leaderless mRNAs by mammalian mitochondrial ribosomes. J Biol Chem 285: 28379-28386. https://doi.org/10.1074/jbc.M110.149054 |
Figures(2)

Chunzheng Li, Chenyu Wei, Gongke Zhao, Xianguang Yang. Cancer cells remodeling and quality control are inextricably linked to autophagy[J]. AIMS Molecular Science, 2023, 10(2): 109-126. doi: 10.3934/molsci.2023009







 DownLoad:
DownLoad: