In this work, we investigated the finite-time passivity problem of neutral-type complex-valued neural networks with time-varying delays. On the basis of the Lyapunov functional, Wirtinger-type inequality technique, and linear matrix inequalities (LMIs) approach, new sufficient conditions were derived to ensure the finite-time boundedness (FTB) and finite-time passivity (FTP) of the concerned network model. At last, two numerical examples with simulations were presented to demonstrate the validity of our criteria.
Citation: Haydar Akca, Chaouki Aouiti, Farid Touati, Changjin Xu. Finite-time passivity of neutral-type complex-valued neural networks with time-varying delays[J]. Mathematical Biosciences and Engineering, 2024, 21(5): 6097-6122. doi: 10.3934/mbe.2024268
In this work, we investigated the finite-time passivity problem of neutral-type complex-valued neural networks with time-varying delays. On the basis of the Lyapunov functional, Wirtinger-type inequality technique, and linear matrix inequalities (LMIs) approach, new sufficient conditions were derived to ensure the finite-time boundedness (FTB) and finite-time passivity (FTP) of the concerned network model. At last, two numerical examples with simulations were presented to demonstrate the validity of our criteria.

| [1] |
C. Aouiti, R. Sakthivel, F. Touati, Global dissipativity of fuzzy bidirectional associative memory neural networks with proportional delays, Iran. J. Fuzzy Syst., 18 (2021), 65–80. https://doi.org/10.22111/ijfs.2021.5914 doi: 10.22111/ijfs.2021.5914

|
| [2] |
F. Wu, T. Kang, Y. Shao, Q. Wang, Stability of Hopfield neural network with resistive and magnetic coupling, Chaos Solitons Fractals, 172 (2023), 113569. https://doi.org/10.1016/j.chaos.2023.113569 doi: 10.1016/j.chaos.2023.113569
|
| [3] |
B. B. He, H. C. Zhou, Asymptotic stability and synchronization of fractional order Hopfield neural networks with unbounded delay, Math. Methods Appl. Sci., 46 (2023), 3157–3175. https://doi.org/10.1002/mma.8000 doi: 10.1002/mma.8000
|
| [4] |
R. P. Agarwal, S. Hristova, Stability of delay Hopfield neural networks with generalized proportional Riemann-Liouville fractional derivative, AIMS Math., 8 (2023), 26801–26820. https://doi.org/10.3934/math.20231372 doi: 10.3934/math.20231372
|
| [5] |
P. Li, R. Gao, C. Xu, J. Shen, S. Ahmad, Y. Li, Exploring the impact of delay on Hopf bifurcation of a type of BAM neural network models concerning three nonidentical delays, Neural Process. Lett., 55 (2023), 1595–11635. https://doi.org/10.1007/s11063-023-11392-0 doi: 10.1007/s11063-023-11392-0
|
| [6] |
B. Zhou, Q. Song, Boundedness and complete stability of complex-valued neural networks with time delay, IEEE Trans. Neural Networks Learn. Syst., 24 (2013), 1227–1238. https://doi.org/10.1109/TNNLS.2013.2247626 doi: 10.1109/TNNLS.2013.2247626
|
| [7] |
Z. Zhang, X. Liu, J. Chen, R. Guo, S. Zhou, Further stability analysis for delayed complex-valued recurrent neural networks, Neurocomputing, 251 (2017), 81–89. https://doi.org/10.1016/j.neucom.2017.04.013 doi: 10.1016/j.neucom.2017.04.013
|
| [8] |
M. S. Ali, G. Narayanan, Z. Orman, V. Shekher, S. Arik, Finite time stability analysis of fractional-order complex-valued memristive neural networks with proportional delays, Neural Process. Lett., 51 (2020), 407–426. https://doi.org/10.1007/s11063-019-10097-7 doi: 10.1007/s11063-019-10097-7
|
| [9] |
Z. Zhang, J. Cao, Periodic solutions for complex-valued neural networks of neutral type by combining graph theory with coincidence degree theory, Adv. Differ. Equations, 2018 (2018), 1–23. https://doi.org/10.1186/s13662-018-1716-6 doi: 10.1186/s13662-018-1716-6
|
| [10] |
Y. Shi, X. Chen, P. Zhu, Dissipativity for a class of quaternion-valued memristor-based neutral-type neural networks with time-varying delays, Math. Methods Appl. Sci., 46 (2023), 18166–18184. https://doi.org/10.1002/mma.9551 doi: 10.1002/mma.9551
|
| [11] |
N. Li, W. X. Zheng, Passivity analysis for quaternion-valued memristor-based neural networks with time-varying delay, IEEE Trans. Neural Networks Learn. Syst., 31 (2019), 639–650. https://doi.org/10.1109/TNNLS.2019.2908755 doi: 10.1109/TNNLS.2019.2908755
|
| [12] |
C. Ge, J. H. Park, C. Hua, C. Shi, Robust passivity analysis for uncertain neural networks with discrete and distributed time-varying delays, Neurocomputing, 364 (2019), 330–337. https://doi.org/10.1016/j.neucom.2019.06.077 doi: 10.1016/j.neucom.2019.06.077
|
| [13] |
S. Chandran, R. Ramachandran, J. Cao, R. P. Agarwal, G. Rajchakit, Passivity analysis for uncertain BAM neural networks with leakage, discrete and distributed delays using novel summation inequality, Int. J. Control Autom. Syst., 17 (2019), 2114–2124. https://doi.org/10.1007/s12555-018-0513-z doi: 10.1007/s12555-018-0513-z
|
| [14] |
M. V. Thuan, D. C. Huong, D. T. Hong, New results on robust finite-time passivity for fractional-order neural networks with uncertainties, Neural Process. Lett., 50 (2019), 1065–1078. https://doi.org/10.1007/s11063-018-9902-9 doi: 10.1007/s11063-018-9902-9
|
| [15] |
J. Han, Finite-time passivity and synchronization for a class of fuzzy inertial complex-valued neural networks with time-varying delays, Axioms, 13 (2024), 39. https://doi.org/10.3390/axioms13010039 https://doi.org/10.3390/axioms13010039 doi: 10.3390/axioms13010039
|
| [16] |
A. Chaouki, F. Touati, Global dissipativity of Clifford-valued multidirectional associative memory neural networks with mixed delays, Comput. Appl. Math., 39 (2020), 1–21. https://doi.org/10.1007/s40314-020-01367-5 doi: 10.1007/s40314-020-01367-5
|
| [17] |
S. Guo, B. Du, Global exponential stability of periodic solution for neutral-type complex-valued neural networks, Discrete Dyn. Nat. Soc., 2016 (2016). https://doi.org/10.1155/2016/1267954 doi: 10.1155/2016/1267954
|
| [18] |
Y. P. Liu, L. H. Zhao, Y. W. Shi, S. Y. Ren, J. L. Wang, Finite-time passivity and synchronisation of complex networks with multiple output couplings, Int. J. Control, 96 (2023), 1470–1490. https://doi.org/10.1080/00207179.2022.2053208 doi: 10.1080/00207179.2022.2053208
|
| [19] |
R. Wei, J. Cao, F. E. Alsaadi, Fixed-time passivity of coupled quaternion-valued neural networks with multiple delayed couplings, Soft Comput., 27 (2023), 8959–8970. https://doi.org/10.1007/s00500-022-07500-2 doi: 10.1007/s00500-022-07500-2
|
| [20] |
X. Chen, Q. Song, Global stability of complex-valued neural networks with both leakage time delay and discrete time delay on time scales, Neurocomputing, 121 (2013), 254–264. https://doi.org/10.1016/j.neucom.2013.04.040 doi: 10.1016/j.neucom.2013.04.040
|
| [21] |
Z. Wang, X. Liu, Exponential stability of impulsive complex-valued neural networks with time delay, Math. Comput. Simul., 156 (2019), 143–157. https://doi.org/10.1016/j.matcom.2018.07.006 doi: 10.1016/j.matcom.2018.07.006
|
| [22] |
M. Chinnamuniyandi, S. Chandran, C. Xu, Fractional order uncertain BAM neural networks with mixed time delays: An existence and Quasi-uniform stability analysis, J. Intell. Fuzzy Syst., 46 (2024), 4291–4313. https://doi.org/10.3233/JIFS-234744 https://doi.org/10.3233/JIFS-234744 doi: 10.3233/JIFS-234744
|
| [23] |
G. Velmurugan, R. Rakkiyappan, S. Lakshmanan, Passivity analysis of memristor-based complex-valued neural networks with time-varying delays, Neural Process. Lett., 42 (2015), 517–540. https://doi.org/10.1007/s11063-014-9371-8 doi: 10.1007/s11063-014-9371-8
|
| [24] |
Z. Zhang, S. Yu, Global asymptotic stability for a class of complex-valued Cohen-Grossberg neural networks with time delays, Neurocomputing, 171 (2016), 1158–1166. https://doi.org/10.1016/j.neucom.2015.07.051 doi: 10.1016/j.neucom.2015.07.051
|
| [25] |
H. Wang, S. Duan, T. Huang, L. Wang, C. Li, Exponential stability of complex-valued memristive recurrent neural networks, IEEE Trans. Neural Networks Learn. Syst., 28 (2016), 766–771. https://doi.org/10.1109/TNNLS.2015.2513001 doi: 10.1109/TNNLS.2015.2513001
|
| [26] |
Z. Zhang, C. Lin, B. Chen, Global stability criterion for delayed complex-valued recurrent neural networks, IEEE Trans. Neural Networks Learn. Syst., 25 (2013), 1704–1708. https://doi.org/10.1109/TNNLS.2013.2288943 doi: 10.1109/TNNLS.2013.2288943
|
| [27] |
J. Pan, X. Liu, W. Xie, Exponential stability of a class of complex-valued neural networks with time-varying delays, Neurocomputing, 164 (2015), 293–299. https://doi.org/10.1016/j.neucom.2015.02.024 doi: 10.1016/j.neucom.2015.02.024
|
| [28] |
W. Qian, S. Cong, T. Li, S. Fei, Improved stability conditions for systems with interval time-varying delay, Int. J. Control Autom. Syst., 10 (2012), 1146–1152. https://doi.org/10.1007/s12555-012-0609-9 doi: 10.1007/s12555-012-0609-9
|
| [29] |
J. Hu, J. Wang, Global stability of complex-valued recurrent neural networks with time-delays, IEEE Trans. Neural Networks Learn. Syst., 23 (2012), 853–865. https://doi.org/10.1109/TNNLS.2012.2195028 doi: 10.1109/TNNLS.2012.2195028
|
| [30] |
T. Dong, X. Liao, A. Wang, Stability and Hopf bifurcation of a complex-valued neural network with two time delays, Nonlinear Dyn., 82 (2015), 173–184. https://doi.org/10.1007/s11071-015-2147-5 doi: 10.1007/s11071-015-2147-5
|
| [31] |
X. Xu, J. Zhang, J. Shi, Dynamical behaviour analysis of delayed complex-valued neural networks with impulsive effect, Int. J. Syst. Sci., 48 (2017), 686–694. https://doi.org/10.1080/00207721.2016.1206988 doi: 10.1080/00207721.2016.1206988
|
| [32] |
R. Samidurai, S. Rajavel, J. Cao, A. Alsaedi, F. Alsaadi, B. Ahmad, Delay-partitioning approach to stability analysis of state estimation for neutral-type neural networks with both time-varying delays and leakage term via sampled-data control, Int. J. Syst. Sci., 48 (2017), 1752–1765. https://doi.org/10.1080/00207721.2017.1282060 doi: 10.1080/00207721.2017.1282060
|
| [33] |
M. S. Ali, S. Saravanan, Q. Zhu, Finite-time stability of neutral-type neural networks with random time-varying delays, Int. J. Syst. Sci., 48 (2017), 3279–3295. https://doi.org/10.1080/00207721.2017.1367434 doi: 10.1080/00207721.2017.1367434
|
| [34] |
C. Hua, Y. Wang, S. Wu, Stability analysis of neural networks with time-varying delay using a new augmented Lyapunov—Krasovskii functional, Neurocomputing, 332 (2019), 1–9. https://doi.org/10.1016/j.neucom.2018.08.044 doi: 10.1016/j.neucom.2018.08.044
|
| [35] |
I. Khonchaiyaphum, N. Samorn, T. Botmart, K. Mukdasai, Finite-time passivity analysis of neutral-type neural networks with mixed time-varying delays, Mathematics, 9 (2021), 3321. https://doi.org/10.3390/math9243321 doi: 10.3390/math9243321
|
| [36] |
J. Xiao, Z. Zeng, Finite-time passivity of neural networks with time varying delay, J. Franklin Inst., 357 (2020), 2437–2456. https://doi.org/10.1016/j.jfranklin.2020.01.023 doi: 10.1016/j.jfranklin.2020.01.023
|
| [37] |
S. Saravanan, M. S. Ali, A. Alsaedi, B. Ahmad, Finite-time passivity for neutral-type neural networks with time-varying delays—via auxiliary function-based integral inequalities, Nonlinear Anal. Modell. Control, 25 (2020), 206–224. https://doi.org/10.15388/namc.2020.25.16513 doi: 10.15388/namc.2020.25.16513
|
| [38] |
A. Seuret, F. Gouaisbaut, On the use of the Wirtinger inequalities for time-delay systems, IFAC Proc. Vol., 45 (2012), 260–265. https://doi.org/10.3182/20120622-3-US-4021.00035 doi: 10.3182/20120622-3-US-4021.00035
|
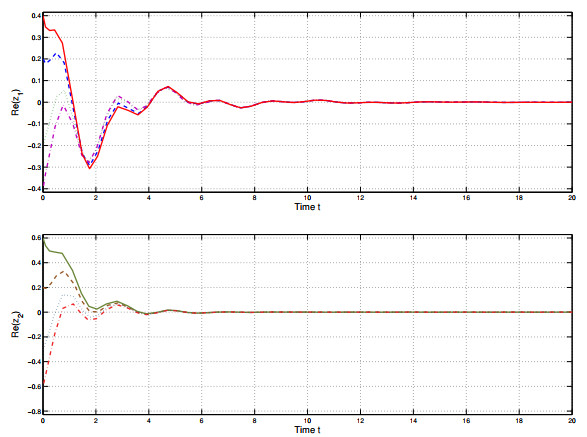





Figures(6) / Tables(1)

Haydar Akca, Chaouki Aouiti, Farid Touati, Changjin Xu. Finite-time passivity of neutral-type complex-valued neural networks with time-varying delays[J]. Mathematical Biosciences and Engineering, 2024, 21(5): 6097-6122. doi: 10.3934/mbe.2024268







 DownLoad:
DownLoad: