The loss and degradation of habitat, Allee effects, climate change, deforestation, hunting-overfishing and human disturbances are alarming and significant threats to the extinction of many species in ecology. When populations compete for natural resources, food supply and habitat, survival to extinction and various other issues are visible. This paper investigates the competition of two species in a heterogeneous environment that are subject to the effect of harvesting. The most realistic harvesting case is connected with the intrinsic growth rate, and the harvesting functions are developed based on this clause instead of random choice. We prove the existence and uniqueness of the solution to the model. Theoretically, we state that, when species coexist, one may drive the other to die out, so both species become extinct, considering all possible rational values of parameters. These results highlight a worthy-of attention study between two populations based on harvesting coefficients. Finally, we solve the model for two spatial dimensions by using a backward Euler, decoupled and linearized time-stepping fully discrete algorithm in a series of examples and observe a match between the theoretical and numerical findings.
Citation: Md. Mashih Ibn Yasin Adan, Md. Kamrujjaman, Md. Mamun Molla, Muhammad Mohebujjaman, Clarisa Buenrostro. Interplay of harvesting and the growth rate for spatially diversified populations and the testing of a decoupled scheme[J]. Mathematical Biosciences and Engineering, 2023, 20(4): 6374-6399. doi: 10.3934/mbe.2023276
The loss and degradation of habitat, Allee effects, climate change, deforestation, hunting-overfishing and human disturbances are alarming and significant threats to the extinction of many species in ecology. When populations compete for natural resources, food supply and habitat, survival to extinction and various other issues are visible. This paper investigates the competition of two species in a heterogeneous environment that are subject to the effect of harvesting. The most realistic harvesting case is connected with the intrinsic growth rate, and the harvesting functions are developed based on this clause instead of random choice. We prove the existence and uniqueness of the solution to the model. Theoretically, we state that, when species coexist, one may drive the other to die out, so both species become extinct, considering all possible rational values of parameters. These results highlight a worthy-of attention study between two populations based on harvesting coefficients. Finally, we solve the model for two spatial dimensions by using a backward Euler, decoupled and linearized time-stepping fully discrete algorithm in a series of examples and observe a match between the theoretical and numerical findings.

| [1] |
G. Dai, M. Tang, Coexistence region and global dynamics of harvested predator-prey system, SIAM J. Appl. Math. 58 (1998), 193–210. https://doi.org/10.1137/S0036139994275799 doi: 10.1137/S0036139994275799

|
| [2] |
E. Braverman, M. Kamrujjaman, Lotka systems with directed dispersal dynamics: Competition and influence of diffusion strategies, Math. Biosci., 279 (2016), 1–12. https://doi.org/10.1016/j.mbs.2016.06.007 doi: 10.1016/j.mbs.2016.06.007
|
| [3] |
F. Brauer, A. C. Soudack, On constant effort harvesting and stocking in a class of predator-prey systems, J. Theor. Biol., 95 (1982), 247–252. https://doi.org/10.1016/0022-5193(82)90242-9 doi: 10.1016/0022-5193(82)90242-9
|
| [4] |
R. D. Parshad, S. Bhowmick, E. Quansah, A. Basheer, R. K. Upadhyay, Predator interference effects on biological control: The "paradox" of the generalist predator revisited, Commun. Nonlinear Sci. Numer. Simul., 39 (2016), 169–184. https://doi.org/10.1016/j.cnsns.2016.02.021 doi: 10.1016/j.cnsns.2016.02.021
|
| [5] |
M. Yavuz, N. Sene, Stability analysis and numerical computation of the fractional predator–prey model with the harvesting rate, Fractal Fract., 4 (2020), 35. https://doi.org/10.3390/fractalfract4030035 doi: 10.3390/fractalfract4030035
|
| [6] |
N. H. Gazi, M. Bandyopadhyay, Effect of time delay on a harvested predator-prey model, J. Appl. Math. Comput., 26 (2008), 263–280. https://doi.org/10.1007/s12190-007-0015-2 doi: 10.1007/s12190-007-0015-2
|
| [7] |
Y. Lou, W. Ni, Diffusion, self-diffusion and cross-diffusion, J. Differ. Equations, 131 (1996), 79–131. https://doi.org/10.1006/jdeq.1996.0157 doi: 10.1006/jdeq.1996.0157
|
| [8] |
M. Kamrujjaman, A. Ahmed, S. Ahmed, Competitive reaction-diffusion systems: Travelling waves and numerical solutions, Adv. Res., 19 (2019), 1–12. https://doi.org/10.9734/air/2019/v19i630140 doi: 10.9734/air/2019/v19i630140
|
| [9] |
B. Wang, Z. Zhang, Dynamics of a diffusive competition model in spatially heterogeneous environment, J. Math. Anal. Appl., 470 (2019), 169–185. https://doi.org/10.1016/j.jmaa.2018.09.062 doi: 10.1016/j.jmaa.2018.09.062
|
| [10] | M. Mohebujjaman, C. Buenrostro, M. Kamrujjaman, T. Khan, Decoupled algorithms for non-linearly coupled reaction-diffusion competition model with harvesting and Stocking, preprint, arXiv: 2209.14144. |
| [11] |
V. Hutson, Y. Lou, K. Mischaikow, Spatial heterogeneity of resources versus Lotka-Volterra dynamics, J. Diff. Equations, 185 (2002), 97–136. https://doi.org/10.1006/jdeq.2001.4157 doi: 10.1006/jdeq.2001.4157
|
| [12] |
Y. Kuang, E. Beretta, Global qualitative analysis of a ratio-dependent predator–prey system, J. Math. Biol., 36 (1998), 389–406. https://doi.org/10.1007/s002850050105 doi: 10.1007/s002850050105
|
| [13] |
L. Korobenko, M. Kamrujjaman, E. Braverman, Persistence and extinction in spatial models with a carrying capacity driven diffusion and harvesting, J. Math. Anal. Appl., 399 (2013), 352–368. https://doi.org/10.1016/j.jmaa.2012.09.057 doi: 10.1016/j.jmaa.2012.09.057
|
| [14] |
K. Y. Lam, W. M. Ni, Uniqueness and complete dynamics in heterogeneous competition-diffusion systems, SIAM J. Appl. Math., 72 (2012), 1695–1712. https://doi.org/10.1137/120869481 doi: 10.1137/120869481
|
| [15] |
E. Braverman, I. Ilmer, On the interplay of harvesting and various diffusion strategies for spatially heterogeneous populations, J. Theor. Biol., 466 (2019), 106–118. https://doi.org/10.1016/j.jtbi.2019.01.024 doi: 10.1016/j.jtbi.2019.01.024
|
| [16] |
L. Bai, K. Wang, Gilpin–Ayala model with spatial diffusion and its optimal harvesting policy, Appl. Math. Comput., 171 (2005), 531–546. https://doi.org/10.1016/j.amc.2005.01.068 doi: 10.1016/j.amc.2005.01.068
|
| [17] |
I. Zahan, M. Kamrujjaman, S. Tanveer, Mathematical study of a resource based diffusion model with Gilpin-Ayala growth and harvesting, Bull. Math. Biol., 84 (2022), 120. https://doi.org/10.1007/s11538-022-01074-8 doi: 10.1007/s11538-022-01074-8
|
| [18] |
A. Martin, S. Ruan, Predator-prey models with delay and prey harvesting, J. Math. Biol. 43 (2001), 247–267. https://doi.org/10.1007/s002850100095 doi: 10.1007/s002850100095
|
| [19] | R. S. Cantrell, C. Cosner, Spatial Ecology via Reaction-Diffusion Equations, Wiley, Hoboken, 2004. |
| [20] |
E. Braverman, M. Kamrujjaman, L. Korobenko, Competitive spatially distributed population dynamics models: Does diversity in diffusion strategies promote coexistence?, Math. Biosci., 264 (2015), 63–73. https://doi.org/10.1016/j.mbs.2015.03.004 doi: 10.1016/j.mbs.2015.03.004
|
| [21] |
L. Korobenko, E. Braverman, On evolutionary stability of carrying capacity driven dispersal in competition with regularly diffusing populations, J. Math. Biol., 69 (2014), 1181–1206. https://doi.org/10.1007/s00285-013-0729-8 doi: 10.1007/s00285-013-0729-8
|
| [22] |
L. Korobenko, E. Braverman, On logistic models with a carrying capacity dependent diffusion: Stability of equilibria and coexistence with a regularly diffusing population, Nonlinear Anal.: Real World Appl., 13 (2012), 2648–2658. https://doi.org/10.1016/j.nonrwa.2011.12.027 doi: 10.1016/j.nonrwa.2011.12.027
|
| [23] | C. V. Pao, Nonlinear Parabolic and Elliptic Equations, Springer, New York, 1992. |
| [24] | S. B. Hsu, H. L. Smith, P. Waltman, Competitive exclusion and coexistence for competitive systems on ordered Banach spaces, Trans. Am. Math. Soc., 348 (1996) 4083–4094. |
| [25] |
E. Braverman, M. Kamrujjaman, Competitive–cooperative models with various diffusion strategies, Comput. Math. Appl., 72 (2016), 653–662. https://doi.org/10.1016/j.camwa.2016.05.017 doi: 10.1016/j.camwa.2016.05.017
|
| [26] |
F. Hecht, New development in FreeFem++, J. Numer. Math., 20 (2012), 251–266. http://dx.doi.org/10.1515/jnum-2012-0013 doi: 10.1515/jnum-2012-0013
|
| [27] |
D. Jiang, K. Lam, Y. Lou, Z. Wang, Monotonicity and global dynamics of a nonlocal two-species phytoplankton model, SIAM J. Appl. Math., 79 (2019), 716–742. https://doi.org/10.1137/18M1221588 doi: 10.1137/18M1221588
|
| [28] |
M. Kamrujjaman, Dispersal dynamics: Competitive symbiotic and predator-prey interactions, J. Adv. Math. Appl., 6 (2017), 7–17. http://dx.doi.org/10.1166/jama.2017.1122 doi: 10.1166/jama.2017.1122
|
| [29] |
M. Kamrujjaman, Directed vs regular diffusion strategy: Evolutionary stability analysis of a competition model and an ideal free pair, Differ. Equation Appl., 11 (2019), 267–290. http://dx.doi.org/10.7153/dea-2019-11-11 doi: 10.7153/dea-2019-11-11
|
| [30] |
M. Kamrujjaman, K. N. Keya, Global analysis of a directed dynamics competition model, J. Adv. Math. Comput. Sci., 27 (2018), 1–14. http://dx.doi.org/10.9734/JAMCS/2018/41247 doi: 10.9734/JAMCS/2018/41247
|
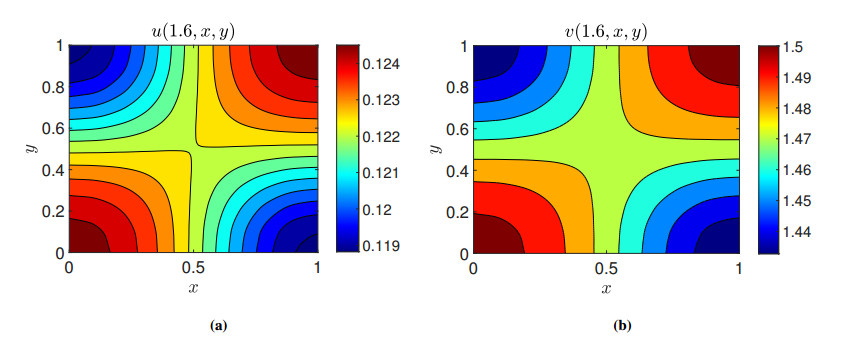
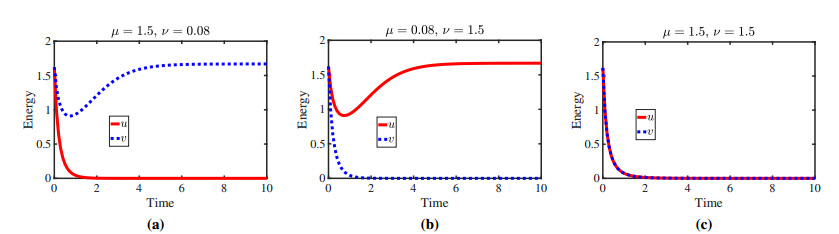
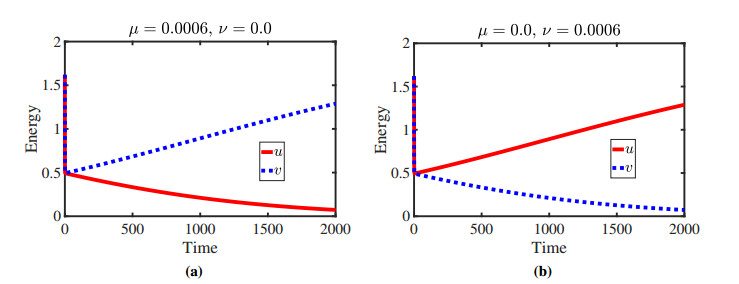
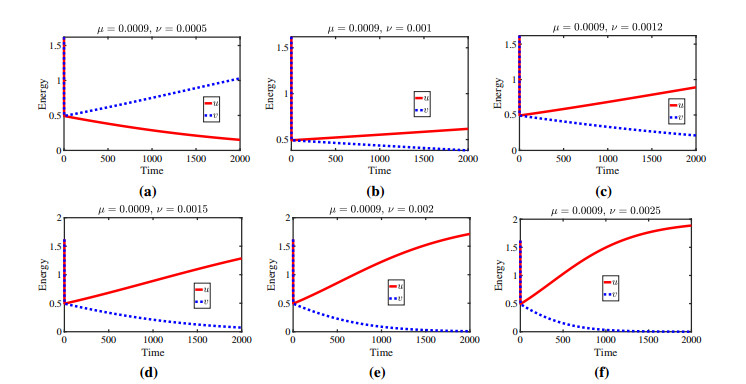
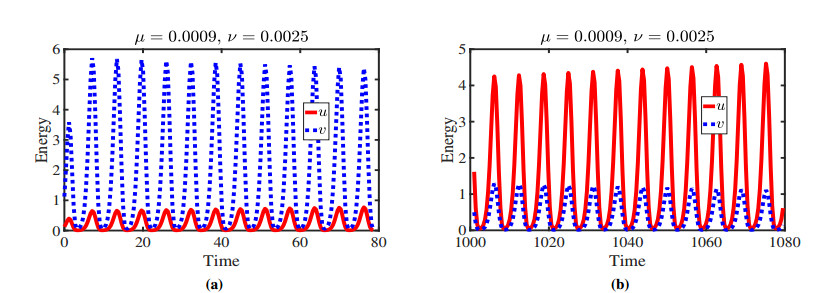
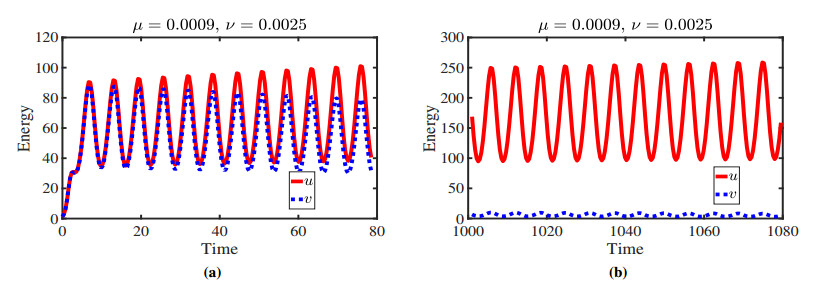
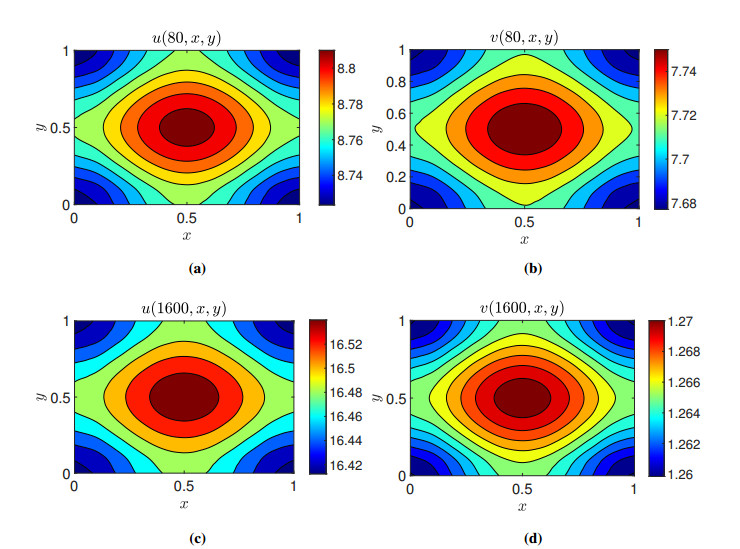
Figures(11)

Md. Mashih Ibn Yasin Adan, Md. Kamrujjaman, Md. Mamun Molla, Muhammad Mohebujjaman, Clarisa Buenrostro. Interplay of harvesting and the growth rate for spatially diversified populations and the testing of a decoupled scheme[J]. Mathematical Biosciences and Engineering, 2023, 20(4): 6374-6399. doi: 10.3934/mbe.2023276







 DownLoad:
DownLoad: