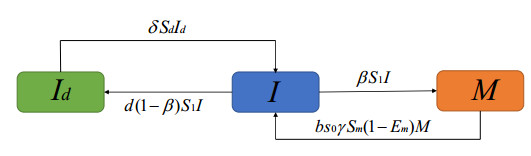
A discrete stage-structured tick population dynamical system with diapause is studied, and spraying acaricides as the control strategy is considered in detail. We stratify vector populations in terms of their maturity status as immature and mature subgroups. The immature subgroup is divided into two categories: normal immature and diapause immature. We compute the net reproduction number $ R_0 $ and perform a qualitative analysis. When $ R_0 < 1 $, the global asymptotic stability of tick-free fixed point is well proved by the inherent projection matrix; there exists a unique coexistence fixed point and the conditions for its asymptotic stability are obtained if and only if $ R_0 > 1; $ the model has transcritical bifurcation if $ R_0 = 1. $ Moreover, we calculate the net reproduction numbers of the model with constant spraying acaricides and periodic spraying acaricides, respectively, and compare the effects of the two methods on controlling tick populations.
Citation: Ning Yu, Xue Zhang. Discrete stage-structured tick population dynamical system with diapause and control[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12981-13006. doi: 10.3934/mbe.2022606
A discrete stage-structured tick population dynamical system with diapause is studied, and spraying acaricides as the control strategy is considered in detail. We stratify vector populations in terms of their maturity status as immature and mature subgroups. The immature subgroup is divided into two categories: normal immature and diapause immature. We compute the net reproduction number $ R_0 $ and perform a qualitative analysis. When $ R_0 < 1 $, the global asymptotic stability of tick-free fixed point is well proved by the inherent projection matrix; there exists a unique coexistence fixed point and the conditions for its asymptotic stability are obtained if and only if $ R_0 > 1; $ the model has transcritical bifurcation if $ R_0 = 1. $ Moreover, we calculate the net reproduction numbers of the model with constant spraying acaricides and periodic spraying acaricides, respectively, and compare the effects of the two methods on controlling tick populations.

| [1] | V. R. Alekseev, B. D. Stasio, J. J. Gilbert, Diapause in Aquatic Invertebrates Theory and Human Use, Springer-Netherlands, New York, 2007. https://doi.org/10.1007/978-1-4020-5680-2_4 |
| [2] | I. Hodek, H. F. van Emden, A. Honěk, Ecology and Behaviour of the Ladybird Beetles (Coccinellidae), Spie Asia-pacific Remote Sensing, International Society for Optics and Photonics, Wiley Online Library, 2012. |
| [3] |
G. Pritchard, The roles of temperature and diapause in the life history of a temperate-zone dragonfly: Argia vivida (Odonata: Coenagrionidae), Eco. Entomol., 14 (2010), 99–108. https://doi.org/10.1111/j.1365-2311.1989.tb00759.x doi: 10.1111/j.1365-2311.1989.tb00759.x

|
| [4] |
H. G. Andrewartha, Diapause in relation to the ecology of insects, Biol. Rev., 27 (1952), 50–-107. https://doi.org/10.1111/j.1469-185x.1952.tb01363.x doi: 10.1111/j.1469-185x.1952.tb01363.x
|
| [5] |
D. F. A. Diniz, C. M. R. de Albuquerque, L. O. Oliva, M. A. V. de Melo-Santos, C. F. J. Ayres, Diapause and quiescence: dormancy mechanisms that contribute to the geographical expansion of mosquitoes and their evolutionary success, Parasit. Vectors, 10 (2017). https://doi.org/10.1186/s13071-017-2235-0 doi: 10.1186/s13071-017-2235-0
|
| [6] |
L. Zhang, Z. C. Wang, Spatial dynamics of a diffusive predator-prey model with stage structure, Discrete Cont. Dyn-B, 20 (2015), 1831–1853. https://doi.org/10.3934/dedsb.2015.20.1831 doi: 10.3934/dedsb.2015.20.1831
|
| [7] |
D. Sadhukhan, B. Mondal, M. Maiti, Discrete age-structured population model with age dependent harvesting and its stability analysis, Appl. Math. Comput., 201 (2008), 631–639. https://doi.org/10.1016/j.amc.2007.12.063 doi: 10.1016/j.amc.2007.12.063
|
| [8] |
K. S. Jatav, J. Dhar, A. K. Nagar, Mathematical study of stage-structured pests control through impulsively released natural enemies with discrete and distributed delays, Appl. Math. Comput., 238 (2014), 511–526. https://doi.org/10.1016/j.amc.2014.04.029 doi: 10.1016/j.amc.2014.04.029
|
| [9] |
J. V. Buskirk, R. S. Ostfeld, Controlling Lyme Disease by Modifying the Density and Species Composition of Tick Hosts, Ecol. Appl., 5 (1995), 1133–1140. https://doi.org/10.2307/2269360 doi: 10.2307/2269360
|
| [10] |
R. S. Ostfeld, F. Keesing, Biodiversity and Disease Risk: the Case of Lyme Disease, Conserv. Biol., 14 (2000), 722–728. https://doi.org/10.1046/j.1523-1739.2000.99014.x doi: 10.1046/j.1523-1739.2000.99014.x
|
| [11] |
R. Rosà, A. Pugliese, Effects of tick population dynamics and host densities on the persistence of tick-borne infections, Math. Biosci., 208 (2007), 216–240. https://doi.org/10.1016/j.mbs.2006.10.002 doi: 10.1016/j.mbs.2006.10.002
|
| [12] |
V. N. Belozerov, L. J. Fourie, D. J. Kok, Photoperiodic Control of Developmental Diapause in Nymphs of Prostriate Ixodid Ticks (Acari: Ixodidae), Exp. Appl. Acarol., 28 (2002), 163–168. https://doi.org/10.1023/A:1025377829119 doi: 10.1023/A:1025377829119
|
| [13] | V. N. Belozerov, Diapause and Biological Rhythms in Ticks, in Physiology of Ticks, Pergamon, 1982,469–500. https://doi.org/10.1016/B978-0-08-024937-7.50018-4 |
| [14] |
J. S. Gray, Mating and behavioural diapause inIxodes ricinus L, Exp. Appl. Acarol., 3 (1987), 61–71. https://doi.org/10.1007/BF01200414 doi: 10.1007/BF01200414
|
| [15] |
Y. J. Lou, K. H. Liu, D. He, D. Gao, S. Ruan, Modelling diapause in mosquito population growth, J. Math. Biol., 78 (2019), 2259–2288. https://doi.org/10.1007/s00285-019-01343-6 doi: 10.1007/s00285-019-01343-6
|
| [16] |
X. Zhang, J. H. Wu, Critical diapause portion for oscillations: Parametric trigonometric functions and their applications for Hopf bifurcation analyses, Math. Methods Appl. Sci., 42 (2019), 1363–1376. https://doi.org/10.1002/mma.5424 doi: 10.1002/mma.5424
|
| [17] |
H. Shu, W. Xu, X. S. Wang, J. Wu, Complex dynamics in a delay differential equation with two delays in tick growth with diapause, J. Differ. Equ., 269 (2020), 10937–10963. https://doi.org/10.1016/j.jde.2020.07.029 doi: 10.1016/j.jde.2020.07.029
|
| [18] |
F. D. Guerrero, R. Miller, A. P. de Leon, Cattle tick vaccines: many candidate antigens, but will a commercially viable product emerge? Int. J. Parasitol., 42 (2012), 421–427. https://doi.org/10.1016/j.ijpara.2012.04.003 doi: 10.1016/j.ijpara.2012.04.003
|
| [19] |
R. Beverton, On the Dynamics of Exploited Fish Populations, Rev. Fish Biol. Fish., 4 (2014), 259–260. https://doi.org/10.1007/BF00044132 doi: 10.1007/BF00044132
|
| [20] | H. Caswell, Matrix Population Models, 2nd edition, Wiley Online Library, Sinauer Sunderland, 2001. https://doi.org/10.1002/9780470057339.vam006m |
| [21] |
J. M. Cushing, Y. Zhou, The net reproduction value and stability in matrix population models, Nat. Resour. Model., 8 (1994), 297–333. https://doi.org/10.1111/j.1939-7445.1994.tb00188.x doi: 10.1111/j.1939-7445.1994.tb00188.x
|
| [22] | J. M. Cushing, An Introduction to Structured Population Dynamics, Society for Industrial and Applied Mathematics, 1998. https://doi.org/10.1137/1.9781611970005.ch3 |
| [23] |
L. Allen, P. Driessche, The basic reproduction number in some discrete- time epidemic models, J. Differ. Equ. Appl., 14 (2018), 1127–1147. https://doi.org/10.1080/10236190802332308 doi: 10.1080/10236190802332308
|
| [24] |
J. K. Hale, P. Waltman, Persistence in infinite-dimensional systems, SIAM J. Math. Anal., 20 (1989), 388–395. https://doi.org/10.1137/0520025 doi: 10.1137/0520025
|
| [25] | J. Carr, Applications of Center Manifold Theory, Springer, New York, 1981. https://doi.org/10.1007/978-1-4612-5929-9 |
| [26] |
L. M. Ladino, J. C. Valverde, Discrete time population dynamics of a two-stage species with recruitment and capture, Chaos Solitons Fractals, 85 (2016), 143–150. https://doi.org/10.1016/j.chaos.2016.01.032 doi: 10.1016/j.chaos.2016.01.032
|
| [27] |
M. Parsamanesh, M. Erfanian, S. Mehrshad, Stability and bifurcations in a discrete-time epidemic model with vaccination and vital dynamics, BMC Bioinformatics, 21 (2020), 1–15. https://doi.org/10.1186/s12859-020-03839-1 doi: 10.1186/s12859-020-03839-1
|
| [28] |
A. S. Ackleh, P. D. Leenheer, Discrete three-stage population model: persistence and global stability results, J. Biol. Dyn., 2 (2008), 415–427. https://doi.org/10.1080/17513750802001812 doi: 10.1080/17513750802001812
|
| [29] | A. D. Barbour, E. R. Lewis, Network Models in Population Biology, Springer, New York, 1997. https://doi.org/10.1007/978-3-642-81134-0 |
| [30] |
J. Hofbauer, W. H. So, Uniform persistence and repellors for maps, Proc. Am. Math. Soc., 107 (1989), 1137–1142. https://doi.org/10.2307/2047679 doi: 10.2307/2047679
|
| [31] |
A. S. Ackleh, A discrete two-stage population model: continuous versus seasonal reproduction, J. Differ. Equ. Appl., 13 (2007), 261–274. https://doi.org/10.1080/10236190601079217 doi: 10.1080/10236190601079217
|
| [32] |
S. E. Randolph, Tick ecology: processes and patterns behind the epidemiological risk posed by ixodid ticks as vectors, Parasitol., 129 (2004), 37–65. https://doi.org/10.1017/S0031182004004925 doi: 10.1017/S0031182004004925
|
| [33] |
J. M. Dunn, S. Davis, A. Stacey, M. A. Diuk-Wasser, A simple model for the establishment of tick-borne pathogens of Ixodes scapularis: A global sensitivity analysis of $R_0$, J. Theor. Biol., 335 (2013), 213–221. https://doi.org/10.1016/j.jtbi.2013.06.035 doi: 10.1016/j.jtbi.2013.06.035
|
| [34] |
S. E. Randolph, D. J. Rogers, A generic population model for the African tick Rhipicephalus appendiculatus, Parasitol., 115 (1997), 265–279. https://doi.org/10.1017/S0031182097001315 doi: 10.1017/S0031182097001315
|
| [35] |
P. A. Hancock, R. AcKley, S. Palmer, Modelling the effect of temperature variation on the seasonal dynamics of Ixodes ricinus tick populations, Int. J. Parasitol., 41 (2011), 513–522. https://doi.org/10.1016/j.ijpara.2010.12.012 doi: 10.1016/j.ijpara.2010.12.012
|
| [36] |
V. N. Belozerov, R. L. Naumov, Nymphal diapause and its photoperiodic control in the tick Ixodes scapularis (Acari: Ixodidae), Folia Parasitol., 49 (2002), 314–318. https://doi.org/10.14411/fp.2002.058 doi: 10.14411/fp.2002.058
|
| [37] |
K. P. Shyma, J. P. Gupta, S. Ghosh, Acaricidal effect of herbal extracts against cattle tick Rhipicephalus (Boophilus) microplus using in vitro studies, Parasitol. Res., 113 (2014), 1919-1926. https://doi.org/10.1007/s00436-014-3839-3 doi: 10.1007/s00436-014-3839-3
|
| [38] |
A. Santiago, L. L. Duarte, T. F. Martins, Occurrence of autogeny in a population of Ornithodoros fonsecai (Acari: Argasidae), Ticks Tick Borne Dis., 10 (2019), 1078–1084. https://doi.org/10.1016/j.ttbdis.2019.05.014 doi: 10.1016/j.ttbdis.2019.05.014
|
| [39] |
X. D. Li, S. J. Song, J. H. Wu, Exponential Stability of Nonlinear Systems With Delayed Impulses and Applications, IEEE Trans. Autom. Control, 64 (2019), 4024–4034. https://doi.org/10.1109/TAC.2019.2905271 doi: 10.1109/TAC.2019.2905271
|
| [40] |
X. D. Li, D. W. C. Ho, J. D. Cao, Finite-time stability and settling-time estimation of nonlinear impulsive systems, Automatica, 99 (2019), 361–368. https://doi.org/10.1016/j.automatica.2018.10.024 doi: 10.1016/j.automatica.2018.10.024
|








Figures(9) / Tables(2)

Ning Yu, Xue Zhang. Discrete stage-structured tick population dynamical system with diapause and control[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12981-13006. doi: 10.3934/mbe.2022606







 DownLoad:
DownLoad: